盐度应激下刺参4个转运相关基因的适应表达研究
蒋亚男,田燚,李晓雨,商艳鹏,常亚青
(大连海洋大学 农业农村部北方海水增养殖重点实验室,辽宁 大连 116023)
摘要:为研究急性低盐18 胁迫对刺参Apostichopus japonicus盐度调节相关基因表达的影响,在实验室条件下,以盐度30 为对照组,72 h 为1 个盐度波动周期,采用荧光定量的方法进行了刺参(16.93 g±3.08 g)不同组织不同时间段单羧酸转运蛋白家族16a13(Monocarboxylate Transporters13,SLC16a13)基因、单羧酸转运蛋白家族6a8(Solute Carrier Family6 Member8,SLC6a8)基因、甲壳素受体蛋白(Fibrinogen C Domain-Containing Protein1,FIBCD1)基因和AMPA型谷氨酸受体1(Glutamate Ionotropic Receptor AMPA Type Subunit1,Gria1)基因4个与盐度调节有关的基因差异表达分析。结果表明:SLC16a13、SLC6a8、FIBCD1、Gria1基因均在刺参不同组织中检测到且有不同程度的表达,这4个基因在刺参体腔液、肠、呼吸树中均有表达;在6、12、48 h时,刺参体腔液中SLC16a13基因的表达量均显著高于对照组(P<0.05),在肠组织中,该基因的表达量在1.5、3 h时与对照组无显著性差异(P>0.05),在其余时间点均显著高于对照组(P<0.05),在72 h时表达量达到最高,在呼吸树组织中,该基因表达量在各时间点均显著上调(P<0.05),在72 h表达量达到最高;试验过程中,SLC6a8基因在体腔液中的表达量显著高于其他组织(P<0.05);总体上FIBCD1基因在体腔液中表达最高,在呼吸树中次之,在肠中最低;总体上Gria1基因在肠组织中表达最高,在体腔液中次之,在呼吸树中最低。本研究结果为刺参对环境的适应机制研究提供了数据资料。
关键词: 刺参;低盐胁迫;荧光定量;基因表达
刺参Apostichopus japonicus因食用价值和药用价值均较高,已在中国得到广泛养殖,是中国北方重要的海水养殖品种,具有重要的经济效益和社会效益。中国北方多用池塘养殖模式养殖刺参,雨季、潮汐变化及冰冻都会导致海水盐度急剧变化,江河入海口附近,雨季易受到江河径流的影响,池塘中的海水可能数日处于低盐状态[1],盐度变化对刺参的存活和生长发育具有重要影响,也是对刺参养殖产业造成较大威胁甚至造成重大经济损失的因素之一[2]。
刺参的盐度耐受范围较小,极限生长盐度范围为15~40,适宜生长的盐度为20~35,最适生长盐度为25~30。龚海滨等[3]研究发现,刺参对低盐度和高盐度海水极度不适应,会导致刺参大量死亡,且在低盐度海水中,其死亡速度快于高盐度海水。张少华等[4]研究表明,盐度骤降会使刺参出现吐肠排脏或个别化皮等应激反应。在海水盐度为18时达到最低耐受限,低盐时间过长,刺参会大量死亡。庚宸帆等[1]研究发现,盐度胁迫对刺参渗透调节能力有显著影响,体腔液渗透压与体腔液Na+、K+、Cl-离子浓度有一定的相关性。Geng等[5]研究了盐度对刺参不同组织结构的影响,发现不同盐度下刺参各器官及组织会发生显著变化,其中肠、管足及纵肌变化最明显。盐度越低,对组织的破坏程度越明显,损伤程度也会更大。
近年来,对水产动物盐度调控相关基因的研究取得了一定的进展。有研究表明,水通道蛋白(AQP)在三疣梭子蟹Portunus trituberculatus[6]、花鳗鲡Anguilla marmorata[7]、萨罗罗非鱼Sarotherodon melanotheron[8]、尼罗罗非鱼Tilapia nilotica[9]、刀鲚Coilia nasus[10]和香港牡蛎Crassostrea hongkongensis[11]等物种中的渗透调节作用已被证实。离子转运酶,如Na+/K+-ATPase、V-H+-ATPase、F-H+-ATPase和碳酸酐酶等在渗透调节器官中起主要作用,可维持机体正常的生理代谢水平,并通过参与主动调节体内的离子含量和细胞体积[12]。NKA基因在高盐和低盐适应过程中均起到维持体内渗透平衡的作用,NKA基因在海水虾[13]和大西洋鳕Gadus morhua[14]、金头鲷Sparus aurata L.[15]、漠斑牙鲆Paralichthys lethostigma[16]、尼罗罗非鱼[17]等硬骨鱼类渗透调节中的作用已被证实。Na+/H+-exchanger蛋白基因在三疣梭子蟹低盐环境下发挥重要的渗透调节功能[18]。胞质碳酸酐酶基因(Cytoplasmic carbonic anhydrase,CAc)在凡纳滨对虾Litopenaeus vannamei和三疣梭子蟹高低盐环境下均响应,而磷脂酰肌醇连(Clycosyl-phophatidylinostitol-linked Carbonic Anhydrase,CAg)只对低盐响应[19]。囊性纤维化跨膜传导调节因子(CFTR)基因在舌齿鲈Dicentrarchus labrax机体对水环境盐度变化的应激反应和调节中发挥作用[20]。免疫球蛋白M (Immunoglobulin M, IgM) 在红鳍东方鲀Takifugu rubripes幼鱼的盐度渗透调节过程中发挥作用[21]。热休克蛋白70(Hsp70)和钠钾氯协同转运蛋白1(Na-K-Clcotransporter 1, NKCC1)基因在黑点青鳉鱼Oryzias dancena[22]、吉丽罗非鱼[23]和红鳍东方鲀[21]幼鱼中参与渗透压调节。其中,NKCC1基因能通过维持细胞较高的钠钾离子渗透梯度,从而提高海水虾在高盐环境的适应能力,而RhoGDI基因可通过调整细胞内离子浓度,使细胞在淡水环境下得以生存,在调控淡水虾的渗透调控中起到重要作用[13]。
AMPK 3个亚基参与凡纳滨对虾低盐渗透胁迫[24]和大菱鲆Scophthalmus maximus盐度渗透胁迫过程[25]。胞内氯离子通道蛋白基因PtCLIC[26]、几丁质酶基因PtChtc[27]在三疣梭子蟹的渗透压调节中发挥作用。Commd1基因(Copper Metabolism Murr 1 Domain)在香港牡蛎发育和渗透压调节方面可能发挥作用[28]。花鲈Lateolabrax maculatus GH/IGF轴可能是通过增加GHRs,进而激活下游IGF-1表达而实现参与低渗调控[29]。
目前,国内外对刺参盐度适应机制相关基因的研究相对较少。DD104基因的表达可能与刺参渗透胁迫有关[30]。有研究表明,Hsp70及Hsp90基因是刺参在盐度胁迫下的重要响应因子[31]。甘氨酸转运蛋白、锌转运蛋白基因、Hsp70和神经乙酰胆碱受体基因参与刺参的低盐调节适应过程[2]。本研究中,选取刺参单羧酸转运蛋白家族16a13(Monocarboxylate Transporters 13,SLC16a13)基因、单羧酸转运蛋白家族6a8(Solute Carrier Family 6 Member 8,SLC6a8)基因、甲壳素受体蛋白(Fibrinogen C Domain-Containing Protein 1,FIBCD1)基因和AMPA型谷氨酸受体1(Glutamate Ionotropic Receptor AMPA Type Subunit 1,Gria1)基因,通过实时荧光定量技术,探究4个基因在刺参体腔液、肠和呼吸树中不同胁迫时间下的表达,以期丰富刺参的生理学理论,为刺参健康养殖提供科学依据。
1 材料与方法
1.1 材料
试验用刺参取自辽宁省大连市瓦房店附近海域,暂养于大连海洋大学北方海水增养殖重点实验室同一养殖池中,水温保持在(16.0±1.0)℃,试验前称量其体质量,刺参的平均湿质量为(16.93±3.08)g。
1.2 方法
1.2.1 试验设计 将正常养殖条件下(盐度为30,低盐胁迫0 h)的刺参作为对照组,低盐胁迫不同时间段的刺参作为试验组。试验用水为自来水曝气24 h后与天然海水配制成盐度为18的海水。将刺参放入盐度为18的海水中进行胁迫,并分别在胁迫后1.5、3、6、12、24、48和72 h进行取样,每个时间点随机取3头刺参,分别取对照组及不同胁迫时间点刺参的肠、呼吸树、体腔液,体腔液以3000 r/min离心10 min,获得体腔细胞。最后将各组织保存于超低温冰箱(-80 ℃)中备用。
1.2.2 引物设计与筛选 从刺参被盐度胁迫后的转录组数据库表达谱中筛选出SLC16a13、SLC6a8、FIBCD1和Gria1 4个基因,以β-actin作为内参基因。以上引物均由生工生物工程(上海)股份有限公司合成。试验所用引物见表1。
1.2.3 荧光定量 PCR 总RNA提取后分别检测其浓度和完整性,然后反转录合成cDNA第一条链,之后保存于冰箱(-20 ℃)中,合成的模板等量混合,以减少个体差异,使用SYBR® Premix ExTaqTMII(TaKaRa),在ABI 7500 Real-Time PCR扩增仪上进行实时定量PCR扩增反应。
1.3 数据处理
以正常盐度30为对照组,每一个时间点设置3个重复,对于任意一个样品,计算目标基因和内参基因(β-actin)的△△Ct值,取平均值后再用2-△△Ct法计算基因的相对表达量。用SPSS 19.0软件对数据进行显著性检验,差异显著性水平设为0.05。
表1 荧光定量PCR所用引物
Tab.1 Primers for Real-time PCR used in the experiment
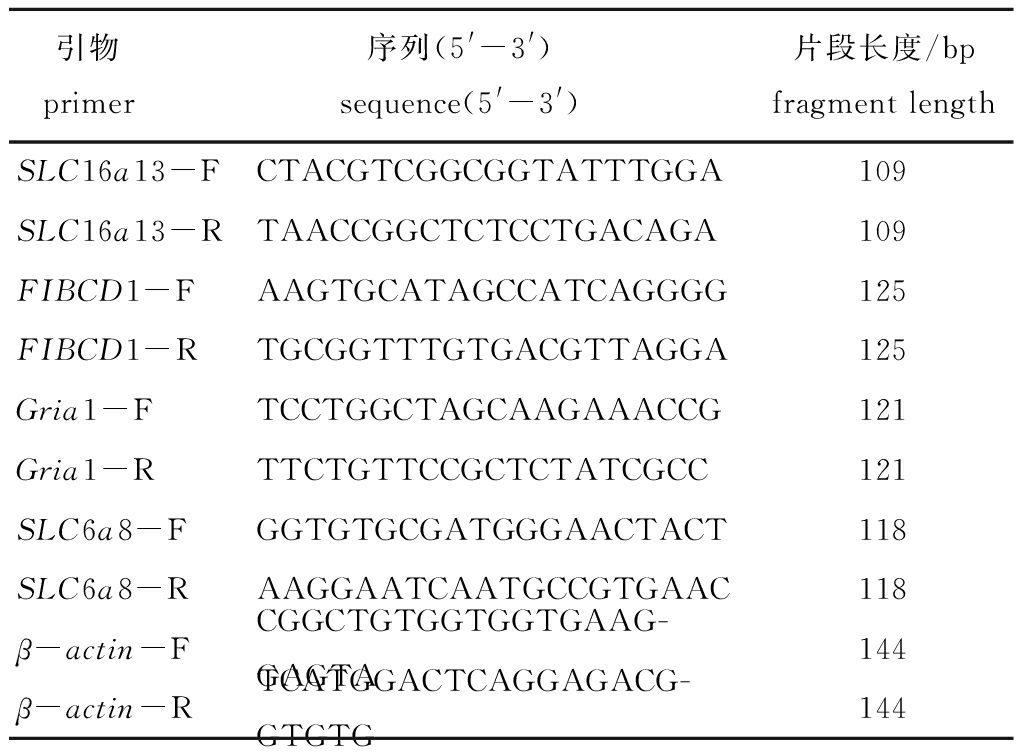
引物 primer 序列(5'-3') sequence(5'-3') 片段长度/bpfragment lengthSLC16a13-FCTACGTCGGCGGTATTTGGA109SLC16a13-RTAACCGGCTCTCCTGACAGA109FIBCD1-FAAGTGCATAGCCATCAGGGG125FIBCD1-RTGCGGTTTGTGACGTTAGGA125Gria1-FTCCTGGCTAGCAAGAAACCG121Gria1-RTTCTGTTCCGCTCTATCGCC121SLC6a8-FGGTGTGCGATGGGAACTACT118SLC6a8-RAAGGAATCAATGCCGTGAAC118β-actin-FCGGCTGTGGTGGTGAAG-GAGTA144β-actin-RTCATGGACTCAGGAGACG-GTGTG144
2 结果与分析
2.1 低盐胁迫下SLC16a13基因在刺参组织中的表达差异
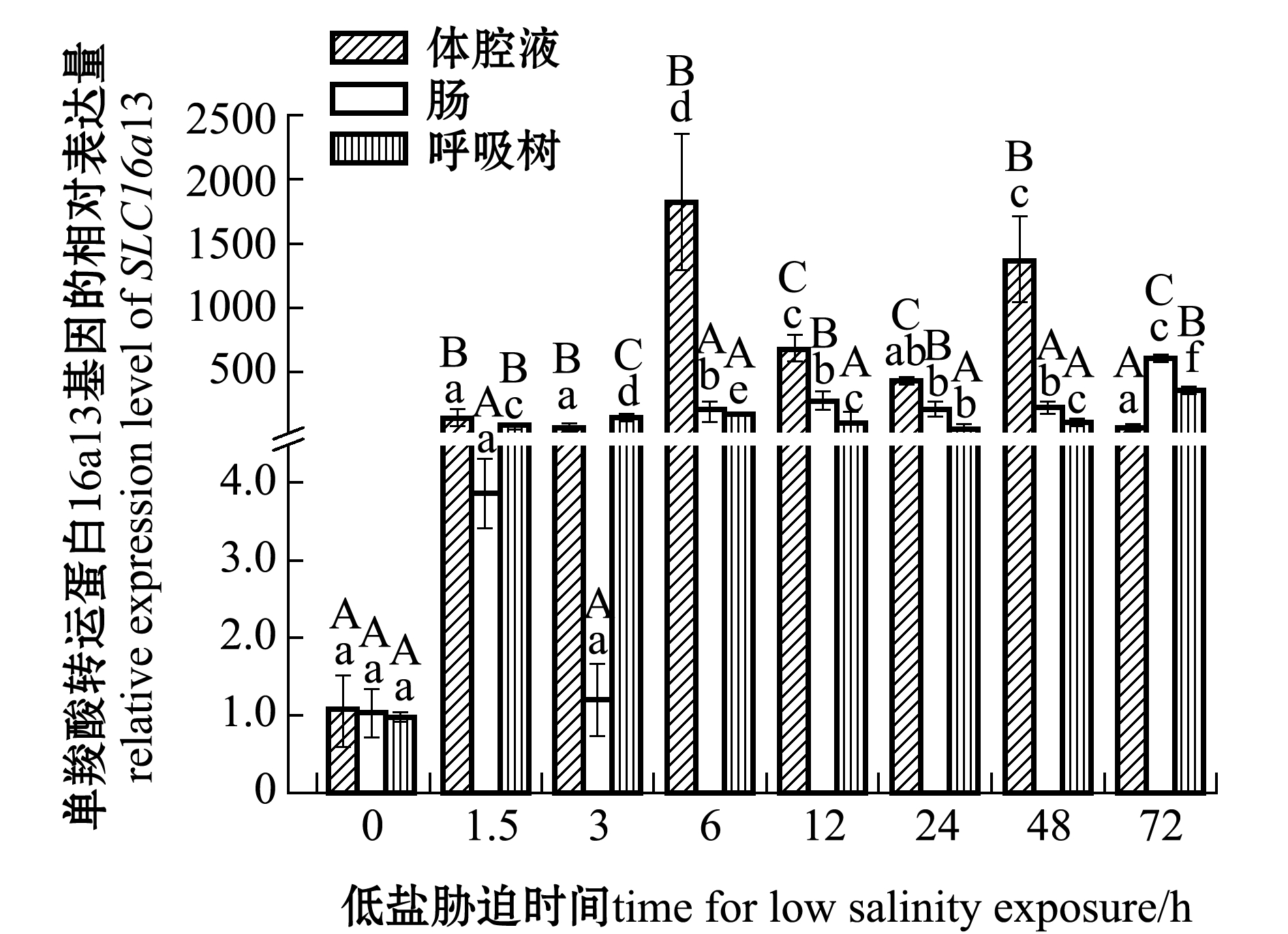
刺参受低盐胁迫后,SLC16a13基因在刺参各组织中的表达情况如图1所示。在体腔液中,与对照组(0 h,盐度为30)相比,SLC16a13基因的表达量在1.5、3、24、72 h时间点无显著性变化(P>0.05),在6、12、48 h时间点表达量显著上调(P<0.05);在肠组织中,SLC16a13基因的表达量在1.5、3 h时间点无显著性变化(P>0.05),在其余时间点表达量均显著上调(P<0.05),在72 h时达到最高;在呼吸树组织中,SLC16a13基因表达量在各时间点均显著上调(P<0.05),在72 h时表达量达到最高。在同一胁迫时间段,1.5、3 h时体腔液和呼吸树中表达量显著高于肠组织(P<0.05),72 h时肠组织中表达量最高,其他时间段SLC16a13在体腔液中表达量均最高,显著高于肠和呼吸树(P<0.05)。
2.2 低盐胁迫下SLC6a8基因在刺参各组织中的表达差异
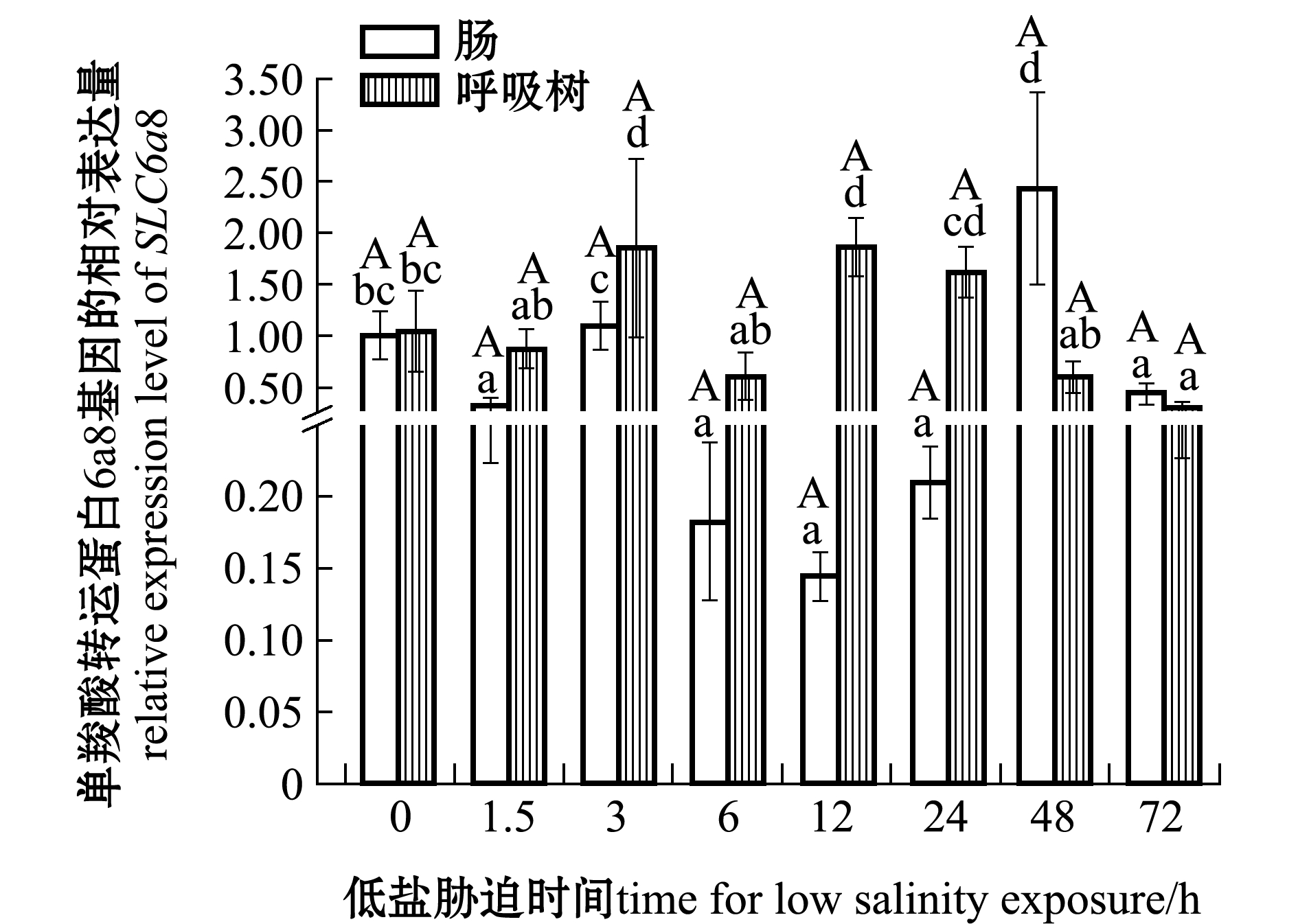
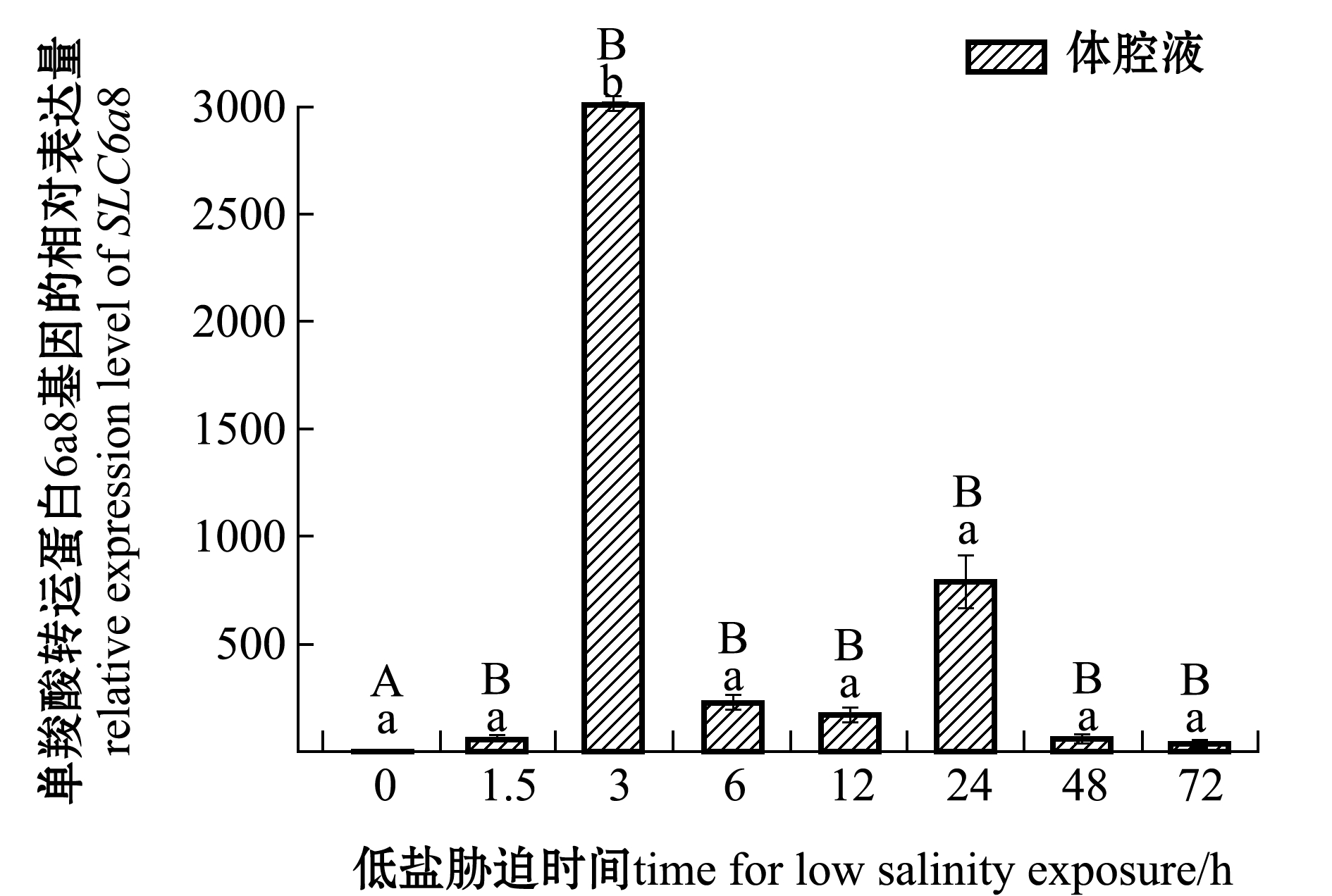
刺参受低盐胁迫后,SLC6a8基因在肠和呼吸树组织中的表达情况如图2所示,在体腔液中的表达情况如图3所示。在肠组织中,与对照组(0 h,盐度30)相比,SLC6a8基因在3、48 h时表达量上调,在其余时间点该基因表达量下调;在呼吸树组织中,在3、12、24 h时该基因表达量上调,12 h时达到最高,在其余时间点表达量下调,72 h时达到最低;在体腔液中,各个时间点该基因表达量均高于对照组,其中在3 h时表达量显著上调(P<0.05),其余时间段表达量均无显著性差异(P>0.05)。在低盐胁迫同一时间段中,体腔液中表达量均显著高于肠和呼吸树(P<0.05)。
2.3 低盐胁迫下FIBCD1基因在刺参组织中的表达差异
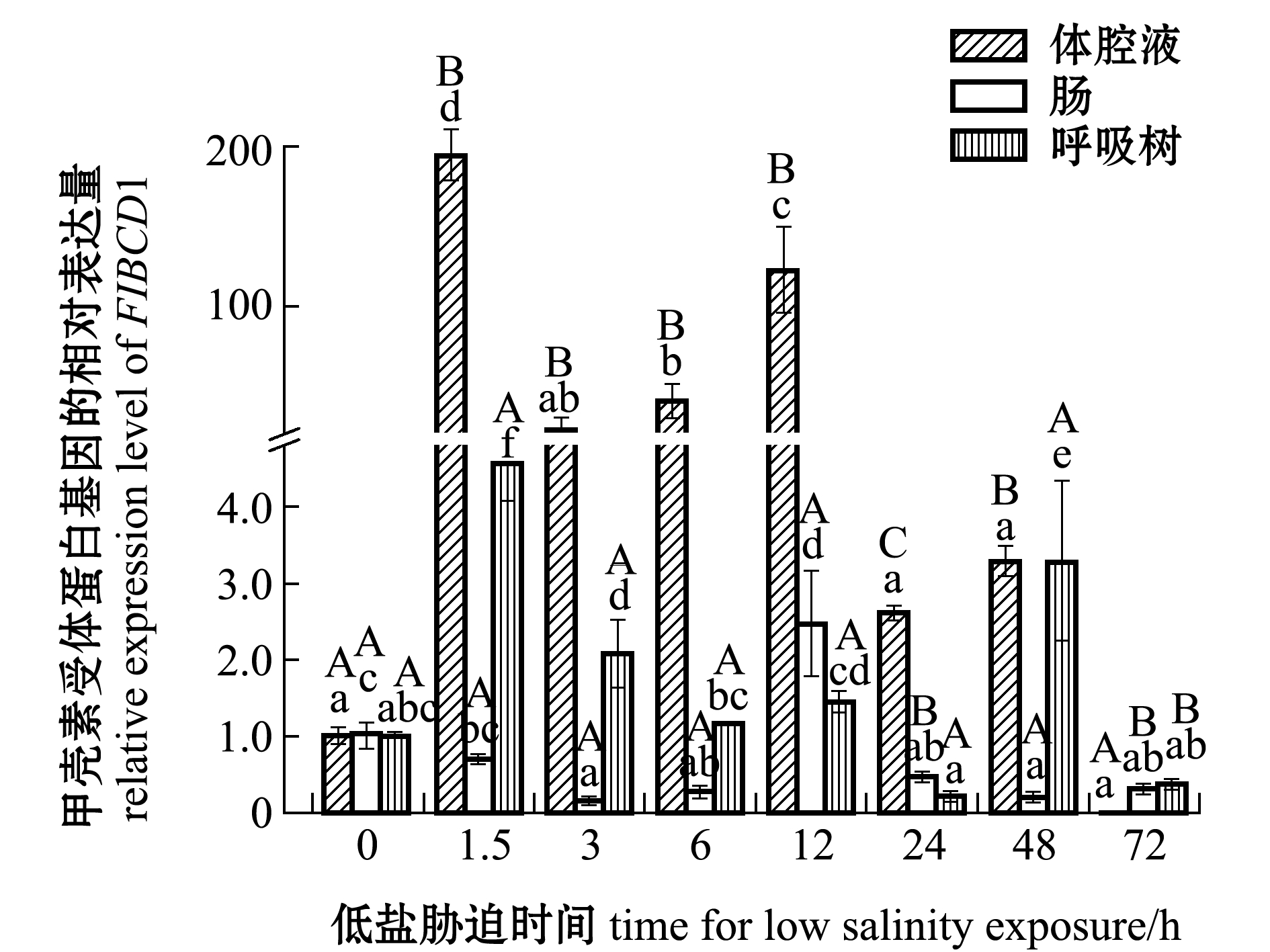
刺参受低盐胁迫后,FIBCD1基因在刺参各组织中的表达情况如图4所示。在体腔液中,与对照组(0 h,盐度30)相比,除72 h时表达量下调外,其余各时间点表达量均上调,在1.5、6、12 h时表达量均显著升高(P<0.05),在其他各时间点该基因表达量无显著性变化(P>0.05)。在肠组织中,与对照组相比,在1.5 h时表达量无显著性变化(P>0.05),在12 h时表达量显著上调(P<0.05),在其余各时间点表达量均显著下调(P<0.05)。在呼吸树组织中,在24、72 h时表达量下调,但与对照组无显著性差异(P>0.05),在其余时间点表达量均有所下调。在同一胁迫时间段,胁迫前期(1.5~24 h)体腔液中表达量显著高于肠和呼吸树(P<0.05),后期72 h时表达量下调至低于肠和呼吸树(P<0.05)。
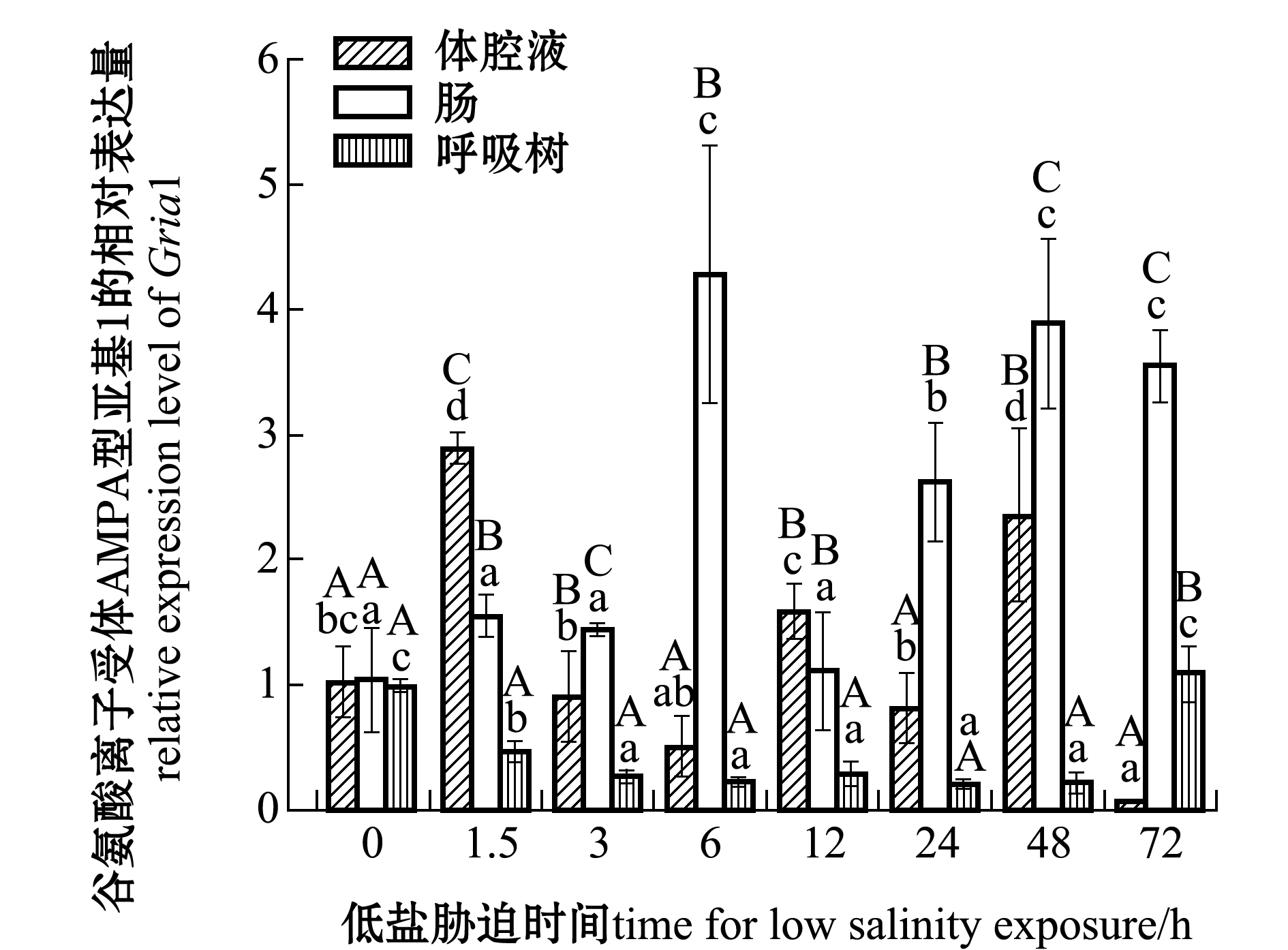
2.4 低盐胁迫下Gria1基因在刺参各组织中的表达差异
刺参受低盐胁迫后,Gria1基因在各组织中的表达情况如图5所示。与对照组(0 h,盐度30)相比,在体腔液中,表达量在3、6、24 h时无显著性差异(P>0.05),在1.5、48 h时显著上调(P<0.05),72 h时显著下调(P<0.05)。在肠组织中,该基因表达量总体趋势为上调,并于6 h时达到最高。在呼吸树组织中,除72 h以外,在其他时间点该基因表达量均下调,且显著低于对照组(P<0.05),72 h时表达量略高于对照组(P>0.05)。同一胁迫时间点,1.5 h时体腔液中Griai的表达量显著高于其他组织(P<0.05),其他时间点(12 h时除外),Gria1在肠组织的表达量均显著高于其他组织(P<0.05)。
3 讨论
3.1 低盐胁迫下单羧酸转运蛋白家族16a13基因的组织表达
单羧酸转运蛋白家族16a13,又称MCT13,属于溶质运载蛋白家族SLC16A亚家族成员,是一类跨膜蛋白,负责转运乙酰乙酸、酮体和乳酸等物质,在维持细胞正常pH值环境和能量代谢方面发挥着重要作用[32]。目前,已发现该家族有14个成员。研究表明,MCTs具有偶联转运细胞新陈代谢中产生的单羧酸与质子的功能[33]。Hirai等[34]研究发现,MCT13在小鼠肝脏和小肠中被PPAR-α激动剂诱导,表明该转运蛋白可能参与营养摄取。Nowik等[35]研究调查小鼠肾脏转录组的变化,检测到SLC16家族的SLC16a13(MCT13)mRNA表达水平有变化。Patel等[36]在研究溶质转运蛋白在重要组织中的作用中多个溶质转运蛋白均表现出上调表达,其中包括MCT13基因在肝脏组织中被上调表达,由此认为多种组织中的各种溶质转运蛋白的稳态表达是精心调控的。本研究结果表明,刺参受低盐胁迫后SLC16a13基因在各组织中总体表现为上调表达,说明该基因参与刺参的盐度适应过程。
3.2 低盐胁迫下单羧酸转运蛋白家族6a8基因的组织表达
单羧酸转运蛋白家族6是通过将溶质耦合到氯与钠转运体,沿着电化学梯度共转运介导溶质穿过质膜[37]。SLC6a8属于单羧酸转运蛋白家族6成员之一,是一种质膜蛋白,其功能是将肌酸转入或者转出细胞,也称为肌酸转运蛋白。其相关途径包括蛋白激酶介导的磷酸化和脱蛋白,具备神经递质和钠同向转运体活性,是摄取肌酸的必需品[38]。肌酸和磷酸肌酸在磷酸盐结合能的储存和传输中起到重要作用,为细胞提供能量[39]。
有研究表明,肌酸在Na+/K +-ATP酶活性、神经递质释放、维持膜电位、Ca2+稳态和离子梯度恢复方面具有重要作用[40]。研究表明,SLC6a8基因在包括骨骼肌、肾脏、小肠、心脏、脑和视网膜在内的各种组织中表达[41]。由于肌酸可逆地结合磷酸盐,可能会补充机体短期内ATP能量的耗竭[42],其细胞的积累能够支持细胞具备储存代谢能量的能力。本研究结果表明,低盐胁迫过程中,SLC6a8基因在肠和呼吸树中的表达总体呈现先上升后下降的趋势,这说明在盐度胁迫过程中,刺参可能需要SLC6a8基因的表达变化来调节刺参机体中肌酸的运输,进而为刺参的盐度适应过程提供能量需求。SLC6a8基因在体腔液的表达一直呈现出上调状态,高表达量说明其在刺参盐度适应过程中发挥重要作用,也说明刺参在盐度适应过程中体腔液对肌酸提供能量的需求大,在体腔液、肠和呼吸树中SLC6a8达到最大表达量的时间点有所不同,说明其在体腔液、肠和呼吸树对肌酸提供能量的需求不同。总之,刺参在盐度胁迫下肌酸转运蛋白基因SLC6a8的表达模式说明,SLC6a8基因可能参与刺参的盐度适应过程,此推测还需要进一步进行试验验证。
3.3 低盐胁迫下谷氨酸离子受体AMPA型亚基1基因的组织表达
谷氨酸离子受体AMPA型亚基1(Gria1/AMPA1)与L-谷氨酸结合后诱导构象改变,导致阳离子通道的打开,进而对Na+、K+有通透性,在中枢神经系统的许多突触中起着重要作用[43]。有研究表明,暴露于急性应激条件下,大鼠的前额皮质椎体神经元AMPA受体调节的突触电流有显著上升趋势[44],谷氨酸受体转运增强,谷氨酸受体在膜上表达升高[45]。在应激刺激下,增加突触传递,突触传递是可逆、迅速的,可能会使突触前膜上的谷氨酸释放增加[46],同时,突触后膜上盐皮质激素受体快速增加,会影响细胞膜AMPA受体的数量,导致海马迅速活跃及神经递质增强。
本研究结果表明,刺参受低盐胁迫后谷氨酸受体蛋白基因Gria1在不同组织和时间点有不同的表达模式,这种结果说明在刺参对盐度的适应过程中,Gria1/AMPA1基因参与其中。
3.4 低盐胁迫下甲壳素受体蛋白基因的组织表达
FIBCD1属于甲壳素受体蛋白家族,其纤维蛋白原结构域可以选择性地与钙依赖的乙酰化组分结合,并且在胞吞作用及免疫防御和免疫反应调制中起到作用[47]。本研究结果表明,刺参受低盐胁迫后FIBCD1基因在各组织中均检测到其表达,说明刺参对盐度的适应过程中可能需要在免疫过程中发挥作用的FIBCD1参与,由于刺参缺乏完善的免疫系统,可能需要免疫相关基因参与其过程,进而达到适应盐度应激的过程。有研究表明,作为重要免疫因子的热休克蛋白基因Hsp70及Hsp90也是刺参在盐度胁迫下的重要响应因子[31]。
综上所述,SLC16a13、SLC6a8、FIBCD1和Gria1 4个基因均参与刺参的盐度适应过程。本研究结果可为刺参的健康养殖提供重要的科学依据,下一步需要进一步验证4个基因在刺参盐度适应过程中发挥的具体功能。
参考文献:
[1] 庚宸帆,田燚,张宇鹏,等.低盐急性胁迫对仿刺参相关生理指标的影响[J].中国水产科学,2015,22(4):666-674.
[2] 傅意然,田燚,常亚青,等.低盐胁迫对刺参4个盐度调节相关基因表达丰度的影响[J].中国水产科学,2014,21(5):902-909.
[3] 龚海滨,王耀兵,邓欢,等.仿刺参对盐度的耐受能力研究[J].水产科学,2009,28(5):284-286.
[4] 张少华,张秀丽,刘振林,等.刺参对盐度的适应范围试验[J].齐鲁渔业,2004,21(12):9-10.
[5] Geng C,Tian Y,Shang f Y,et al.Effect of acute salinity stress on ion homeostasis,Na+/K+-ATPase and histological structure in sea cucumberApostichopus japonicus[J].Springerplus,2016,5:1977.
[6] 王渝,吕建建,刘萍,等.三疣梭子蟹水通道蛋白1 cDNA及其盐度胁迫下的表达分析[J].中国水产科学,2014,21(5):893-901.
[7] 王丹,李多琳,梁奋飞,等.花鳗鲡AQP3基因的分子特征及其应对盐度变化的表达规律[J].海洋渔业,2017,39(5):518-528.
[8] 甘远迪,赵金良,Thammaratsuntorn J,等.萨罗罗非鱼AQP3 cDNA序列克隆及盐度胁迫下组织表达特征[J].动物学杂志,2014,49(4):560-569.
[9] 李茜茜,朱华平,卢迈新,等.盐胁迫对橙色莫桑比克罗非鱼AQP1基因表达的影响[J].基因组学与应用生物学,2016,35(11):2996-3006.
[10] 王美垚,杨健,徐跑,等.刀鲚水通道蛋白1的分子克隆及高盐作用下的表达分析[J].中国水产科学,2017,24(3):449-458.
[11] 万茜,张扬,张跃环,等.香港牡蛎(Crassostrea hongkongensis)水通道蛋白基因AQP1的克隆、分子特性和表达分析[J].海洋与湖沼,2015,46(5):1078-1087.
[12] 张丹,王芳,董双林.周期性盐度波动对凡纳滨对虾游离氨基酸含量及渗透调节相关基因表达的影响[J].中国水产科学,2016,23(5):1130-1136.
[13] 袁剑波.凡纳滨对虾系统发生地位及适应性进化分析[D].青岛:中国科学院海洋研究所,2014.
[14] Larsen P F,Nielsen E E,Meier K,et al.Differences in salinity tolerance and gene expression between two populations of Atlantic cod (Gadus morhua) in response to salinity stress[J].Biochemical Genetics,2012,50(5-6):454-466.
[15] Miguel M J,laiz C R.Osmoregulatory action of PRL,GH,and cortisol in the gilthead seabream (Sparus aurata L.)[J].General and Comparative Endocrinology,2002,129(2):95-103.
[16] Tipsmark C K,Luckenbach J A,Madsen S S,et al.Osmoregulation and expression of ion transport proteins and putative claudins in the gill of southern flounder (Paralichthys lethostigma)[J].Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology,2008,150(3):265-273.
[17] 赵丽慧.盐碱度对尼罗罗非鱼生长、渗透生理及鳃离子转运酶基因表达的影响[D].上海:上海海洋大学,2014.
[18] 马金武,吕建建,刘萍,等.三疣梭子蟹Na+/H+-exchanger基因克隆鉴定及在盐度胁迫下的表达分析[J].水生生物学报,2016,40(5):902-907.
[19] 胡岩岩.凡纳滨对虾和三疣梭子蟹渗透调节相关基因的筛选与表达分析[D].青岛:中国海洋大学,2012.
[20] Bodinier C,Lorin-Nebel C,Charmantier G,et al.Influence of salinity on the localization and expression of the CFTR chloride channel in the ionocytes of juvenileDicentrarchus labrax exposed to seawater and freshwater[J].Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology,2009,153(3):345-351.
[21] 孙梦蕾,吕绘倩,暴宁,等.急性低盐胁迫下红鳍东方鲀幼鱼IgM、NKCC1和Hsp70基因的表达[J].中国水产科学,2017,24(1):65-72.
[22] Kang C K,Tsai H J,Liu C C,et al.Salinity-dependent expression of a Na+,K+,2Cl+cotransporter in gills of the brackish medakaOryzias dancena:a molecular correlate for hyposmoregulatory endurance[J].Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology,2010,157(1):7-18.
[23] 王兵,范武江,李思发.不同盐度下“吉丽”罗非鱼(尼罗罗非鱼♀×萨罗罗非鱼♂)NKCC1a mRNA 的组织特异性表达[J].中国水产科学,2013,18(3):515-522.
[24] Xu C,Li E C,Xu Z X,et al.Molecular characterization and expression of AMP-activated protein kinase in response to low-salinity stress in the Pacific white shrimpLitopenaeus vannamei[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2016,198:79-90.
[25] Zeng L,Liu B,Wu C W,et al.Molecular characterization and expression analysis of AMPKα subunit isoform genes fromScophthalmus maximus responding to salinity stress[J].Fish Physiology and Biochemistry,2016,42(6):1595-1607.
[26] 王渝,吕建建,刘萍,等.三疣梭子蟹(Portunus trituberculatus)胞内氯离子通道蛋白基因克隆及其表达分析[J].海洋与湖沼,2014,45(6):1359-1366.
[27] 张凤,吕建建,刘萍,等.三疣梭子蟹几丁质酶基因克隆鉴定及在低盐胁迫和蜕皮周期中的表达分析[J].海洋与湖沼,2015,46(4):948-957.
[28] 王富轩,肖述,向志明,等.香港牡蛎(Crassostrea hongkongensis)Commd1基因的分子克隆及其在盐度胁迫下的表达分析[J].热带海洋学报,2017,36(1):48-55.
[29] 张沛,迟美丽,温海深,等.盐度调控对花鲈(Lateolabrax maculatus)生长相关基因表达的影响[J].海洋与湖沼,2015,46(2):446-453.
[30] 田燚,莫海波,常亚青.低盐胁迫下仿刺参DD104基因的定量表达分析[J].大连海洋大学学报,2013,28(3):236-240.
[31] 于姗姗,王青林,孟宪亮,等.盐度骤变对仿刺参hsp70及hsp90基因表达的影响[J].中国海洋大学学报,2012,42(9):22-27.
[32] Ålund J.Functional characterization of central transport mechanisms by novel genes[D].Sweden:Uppsala University,2012.
[33] Morris M E,Felmlee M A.Overview of the proton-coupled MCT (SLC16A) family of transporters:characterization,function and role in the transport of the drug of abuseγ-hydroxybutyric acid[J].The AAPS Journal,2008,10(2):311-321.
[34] Hirai T,Fukui Y,Motojima K.PPARalpha agonists positively and negatively regulate the expression of several nutrient/drug transporters in mouse small intestine[J].Biological & Pharmaceutical Bulletin,2007,30(11):2185-2190.
[35] Nowik M,Lecca M R,Velic A,et al.Genome-wide gene expression profiling reveals renal genes regulated during metabolic acidosis[J].Physiological Genomics,2008,32(3):322-334.
[36] Patel O V,Plaut K.Homeorhetic adaptation during the pregnancy to lactation transition:role of solute-linked transporter genes in key metabolic tissues[J].Biology of Reproduction,2011,85(7):556.
[37] Chen N H,Reith M E,Quick M W.Synaptic uptake and beyond:the sodium- and chloride-dependent neurotransmitter transporter family SLC6[J].Pflugers Archiv,2004,447(5):519-531.
[38] Gregor P,Nash S R,Caron M G,et al.Assignment of the creatine transporter gene (SLC6a8) to human chromosome Xq28 telomeric to G6PD[J].Genomics,1995,25(1):332-333.
[39] Wyss M,Kaddurah-Daouk R.Creatine and creatinine metabolism[J].Physiological Reviews,2000,80(3):1107-1213.
[40] Wallimann T,Wyss M,Brdiczka D,et al.Intracellular compartmentation,structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands:the ‘phosphocreatine circuit’for cellular energy homeostasis[J].Biochemical Journal,1992,281:21-40.
[41] Guimbal C,Kilimann M W.A Na+-dependent creatine transporter in rabbit brain,muscle,heart,and kidney.cDNA cloning and functional expression[J].Journalof Biological Chemistry,1993,268(12):8418-8421.
[42] Speer O,Neukomm L J,Murphy R M,et al.Creatine transporters:a reappraisal[J].Molecular and Cellular Biochemistry,2004,256(12):407-424.
[43] Kumar J,Mayer M L.Functional insights from glutamate receptor ion channel structures[J].Annual Review of Physiology,2013,75:313-337.
[44] Groc L,Choquet D,Chaouloff F.The stress hormone corticosterone conditions AMPAR surface trafficking and synaptic potentiation[J].Nature Neuroscience,2008,11(8):868-870.
[45] Yuen E Y,Liu W,Karatsoreos I N,et al.Mechanisms for acute stress-induced enhancement of glutamatergic transmission and working memory[J].Molecular Psychiatry,2011,16(2):156-170.
[46] Olijslagers J E,De Kloet E R,Elgersma Y,et al.Rapid changes in hippocampal CA1 pyramidal cell function via pre- as well as postsynaptic membrane mineralocorticoid receptors[J].European Journal of Neuroscience,2010,27(10):2542-2550.
[47] Schlosser A,Thomsen T,Moeller J B,et al.Characterization of FIBCD1 as an acetyl group-binding receptor that binds chitin[J].The Journal of Immunology,2009,183(6):3800-3809.
Adaptative expression of four transporter-related genes in sea cucumber Apostichopus japonicus exposed to salinity stress
JIANG Ya-nan, TIAN Yi, LI Xiao-yu, SHANG Yan-peng, CHANG Ya-qing
(Key Laboratory of Mariculture & Stock Enhancement in North China’s Sea, Ministry of Agriculture and Rural Affairs, Dalian Ocean University, Dalian 116023, China)
Abstract: The expression profiles of four genes related-salinity regulation were detected in intestine, coelomic fluid, and respiratory tree of sea cucumberApostichopus japonicas with body weight of (16.93±3.08)g reared at a salinity of 30 (as control group) which were abruptly exposed to low salinity of 18 for 1.3, 3,6, 12, 24, 48 and 72 h by RT-PCR, including single carboxylic acid transporter family 16a13 (Monocarboxylate Transporters 13, SLC16a13) gene, single carboxylic acid transporter family 6a8 (Solute Carrier Family 6 member 8, SLC6a8) gene, Chitin receptor protein (Fibrinogen C Domain-Containing Protein 1, FIBCD1) gene, and AMPA type glutamate receptor 1 (Glutamate Ionotropic Receptor AMPA Type Subunit 1, Gria1) gene. The results showed that there was expression of the 4 genes in coelomic fluid, intestine and respiratory tree significantly tree higherSLC16a13 expression level than that in the control group at 6 h, 12 h, and 48 h(P<0.05), except in the intestine at 1.5 h and 3 h, with the maximum at 72 h. In the respiratory tree, the significantly up-regulatedSLC16a13 expression was shown, the maximal expression at 72. During the experiment,SLC16a8 gene was shown to be significantly higher in coelomic fluid that in the other tissues(P<0.05). The maximalFIBCD1 expression level was observed in coelomic fluid, followed by the respiratory tree, and the minimum in the intestine. There was the maximalGria1 expression in the intestine, followed by coelomic fluid and the minimal in the respiratory tree. The findings appear to be the foundation for research of adaptation mechanism of sea cucumber.
Key words: Apostichopus japonicas; low salinity stress; quantitative real-time PCR; gene expression
收稿日期: 2018-01-27
基金项目: 辽宁省教育厅科研项目(L201620);农业农村部北方海水增养殖重点实验室资助项目(2015-MSENC-KF-02)
作者简介: 蒋亚男(1994—), 女, 硕士研究生。E-mail:jiangyn1994@Outlook.com
通信作者: 田燚(1979—), 女, 博士, 副教授。E-mail:tianyi@dlou.edu.cn
中图分类号:S917
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2018.06.003
文章编号:2095-1388(2018)06-0696-07