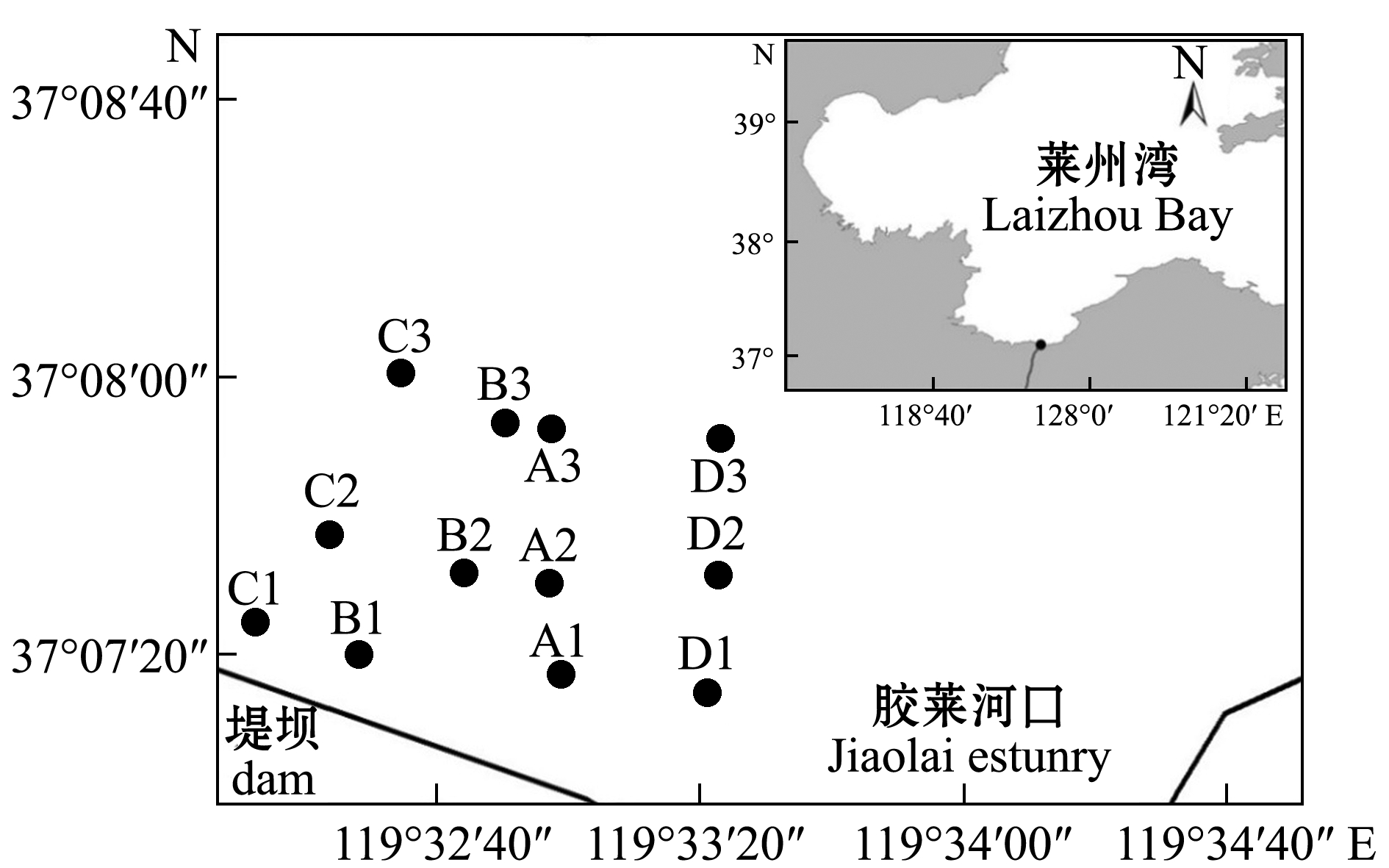
图1 研究区域和调查站点
Fig.1 Study area and sampling stations
李慧峰1、2,杨文波1、2,袁立来1,刘宝祥1,曹坤1,王琳1,丁放1,李慧琴1,李继龙1,李应仁1,黄经献3,张传涛3
(1.中国水产科学研究院资源与环境研究中心,北京 100141;2. 上海海洋大学 海洋科学学院,上海 201306;3. 中国水产科学研究院 下营增殖实验站,山东 昌邑 261300)
摘要:为了解莱州湾南部胶莱河口潮间带大型底栖动物现状,分别于 2016年春季、夏季和秋季对该潮间带大型底栖动物的种类组成、优势种、生物多样性和群落结构特征进行了调查,3次采样共鉴定出大型底栖动物5门28种,其中春季23种,夏季和秋季均为24种。调查结果表明:春、夏、秋3个季节大型底栖动物平均生物量分别为13.77、24.55、18.17 g/m2,3个季节的大型底栖动物平均丰度分别为38.89、68.49、55.07 ind./m2;软体动物为主要优势类群,其次是节肢动物;异白樱蛤Macomaincongrua、红明樱蛤Moerellarutila、托氏虫昌螺Umboniumthomas为春季大型底栖动物优势种;夏季和秋季大型底栖动物优势种相同,依次为异白樱蛤、红明樱蛤、托氏虫昌螺和古氏滩栖螺Batillariacumingi;Shannon-Weiner多样性指数(H′)变化范围为2.13~2.55,Pielou均匀度指数(J)变化范围为0.62~0.73,Margalef物种丰富度指数(D)变化范围为1.17~1.35。研究表明,莱州湾南部胶莱河口潮间带大型底栖动物群落结构存在明显的潮区变化,各季节典型种主要有异白樱蛤、红明樱蛤等,分歧种主要有古氏滩栖螺、大竹蛏Solengrandis、薄壳绿螂Glauconomeprimeana等。
关键词:潮间带;大型底栖动物;时空分布;群落结构
大型底栖动物在潮间带生态系统食物网中有促进食物网内有机质分解、营养物质循环和能量流动的作用,其种类组成和生物量的季节性变动能有效反映生境梯度变化和生态系统功能[1],有助于理解生态系统结构和功能的潜在关系,从而为研究由于气候变化和人类活动引起的生物多样性变化对生态系统的威胁提供重要的线索[2]。同时,大型底栖动物对环境变化敏感[3],某些种类的数量分布对气候变化和环境的变动也具有一定的指示作用,其群落结构的变化往往能够较好地表征潮间带生态系统的稳定性[4],因此,开展大型底栖动物研究对海岸环境监测及沿岸工程环境评价方面具有重要的意义[5]。
莱州湾南部胶莱河入海口处水动力及水文条件变化较大,导致潮间带物质来源复杂,同时,河口处充足的饵料和营养也造就了胶莱河口潮间带大型底栖动物多样性的格局。高昊东等[6]对秋季胶莱河口东岸潮间带底栖生物种类进行了调查,认为潮区间生物组成差别较大,但优势种类相同,认为生物量低与间隙水中高盐、高溴有关。顾炎斌等[7]对东营市河口区春季和秋季不同断面的大型底栖动物种类组成做了调查。但均未见系统地对该水域大型底栖动物群落结构季节和潮区变化进行全面性的时空分析。本研究中,通过对莱州湾胶莱河口潮间带大型底栖动物群落结构特征进行调查,对大型底栖动物种类组成、优势种、丰度和生物量的时空分布和群落结构随潮区变化特征进行了探讨,以期为潮间带生态系统保护利用提供依据,也为本区域大型底栖动物多样性研究和河口区潮间带生态系统的现状评价提供基础资料。
研究区域位于莱州湾南部胶莱河入海处的潮间带区域(119°30′E~119°33′E,37°06′57″N~37°09′45″N)。本研究中根据《海洋调查规范》(GB/T 12763.6—2007)有关原则和要求[8],选取4条大致呈垂直于岸线分布的调查断面A、B、C、D,在选择采样点时包含高潮区(A1、B1、C1、D1)、中潮区(A2、B2、C2、D2)和低潮区(A3、B3、C3、D3),每个断面设置3个调查站位,共设12个站位,每个调查站位重复取样3次,取其平均值(图1)。
图1 研究区域和调查站点
Fig.1 Study area and sampling stations
1.2.1 样品的采集 分别于2016年春季(4月)、夏季(7月)、秋季(10月)进行样品采集,每个小站点采样样方规格为0.25 m×0.25 m×0.30 m,对泥样合并后,现场利用过筛器进行淘洗,并对采集到的大型底栖动物进行固定,最后带回实验室进行种类鉴定、计数和称重。
1.2.2 指标的计算与分析 采用Shannon-Weiner多样性指数(H′)、Margalef种类丰富度指数(D)和Pielou均匀度指数(J)全方位分析群落的多样性,采用Simpson优势度指数(Y)表示优势种类。其计算公式[9]如下:
(1)
D=(S-1)/log2N,
(2)
J=H′/log2S,
(3)
Y=(ni/N)×fi,
(4)
其中:N为所有种类总个体数;S为样品中的种类总数;Pi为第i种的个体数(ni)与总个体数的比值,ni为第i种个体数;fi为第i种出现的站位数占总站位数的比率;Y≥0.02的物种视为群落优势种[10]。
采用聚类分析和非参数多维表序排序(NMDS)分析莱州湾胶莱河口处大型底栖动物的群落结构,分析前先对大型底栖动物的数据进行4次方根标准化。用胁强系数(stress)来衡量NMDS分析结果的优劣,当stress<0.2时,可用二维图表示,图形具有一定的解释意义;当stress<0.1时,可认为是一个好的排序,当stress<0.05时,具有较好的代表性[11-12]。采用单因子相似性分析法(ANOSIM)分析不同潮区间大型底栖动物群落结构差异的显著性,采用相似性百分比(SIMPER)分析各潮区群落结构的相似种和分歧种[13-14]。
采用SPSS 23.0软件和英国普林矛斯海洋研究所开发的PRIMER 5.0软件进行相关数据处理[9]。使用Arcgis 10.3软件绘制站位、丰度和生物量分布图。
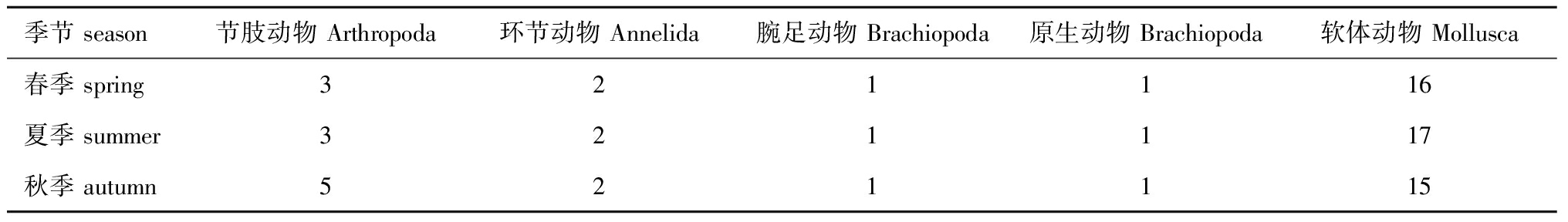
本次调查共鉴定出大型底栖动物5门28种。大型底栖动物在春季出现23种,夏季和秋季均出现24种。其中,软体动物有19种,占大型底栖动物总种数的67.86%;节肢动物5种,占总种类数的17.86%;环节动物2种,占总类数的7.14%;腕足动物和原生动物各1种,均占总种类数的3.57%。从表1可见,春、夏、秋3个季节大型底栖动物门类组成均以软体动物和节肢动物为主。春季,软体动物有16种,占春季总种类数的69.56%,节肢动物3种,占13.04%;夏季,软体动物有17种,占夏季总种类数的70.83%,节肢动物3种,占12.5%;秋季,软体动物有15种,占秋季总种类数的62.5%,节肢动物5种,占20.83%。从季节变化来看,莱州湾南部胶莱河入海口处潮间带夏季大型底栖动物种类数目变化不大,但秋季节肢动物种类数明显增加。
从物种数目在空间上的分布(图2)情况看,采集到物种数沿低潮区(A3、B3、C3、D3)、中潮区(A2、B2、C2、D2)和高潮区(A1、B1、C1、D1)有逐渐增多的趋势。具体表现为低潮区(17种)<中潮区(18种)<高潮区(21种),低潮区、中潮区和高潮区采集到的物种均以软体动物为主,软体动物种类数目占总物种数的比例分别为70.47%、66.67%和63.50%。
表1不同季节大型底栖动物各门类种类数目
Tab.1Numberofspeciesinmacrozoobenthosinvariousphyladifferentseasons
季节 season节肢动物 Arthropoda环节动物 Annelida腕足动物 Brachiopoda原生动物 Brachiopoda软体动物 Mollusca春季 spring321116夏季 summer321117秋季 autumn521115
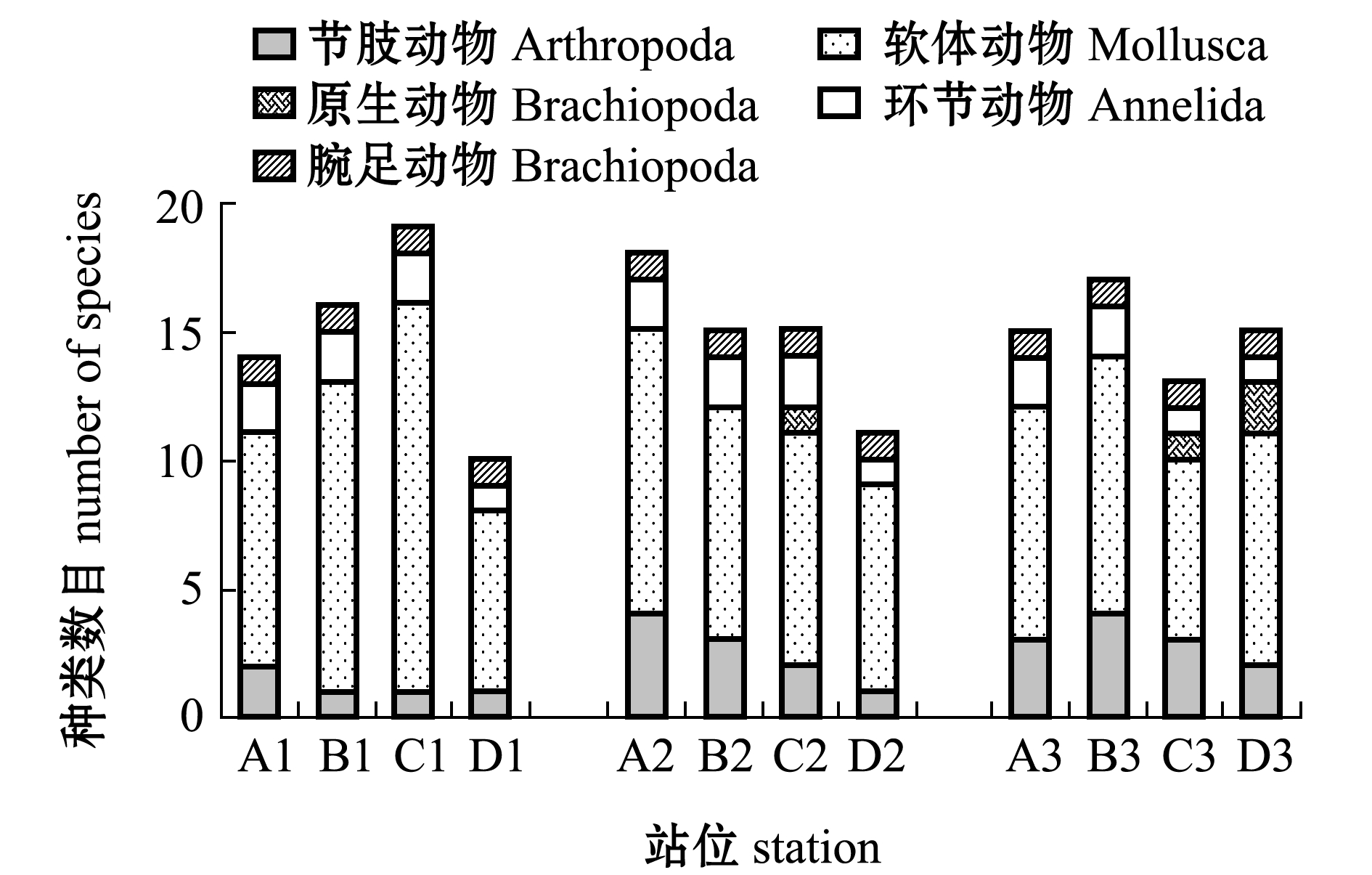
图2 不同站位物种数目
Fig.2 Number of species at different stations
从图3-A可见:大型底栖动物春季的丰度为16~72 ind./m2,最低值区主要出现在D1、C2、B3站位,最高值出现在A2站位,平均丰度为38.89 ind./m2;大型底栖动物夏季的丰度为25.83~167.17 ind./m2,低值区主要出现在D2、A3站位,最高值出现在A1站位,平均丰度为68.49 ind./m2;大型底栖动物秋季的丰度为17.83~89.67 ind./m2,低值区主要出现在C2、C3和D2站位,最高值出现在A2站位,平均丰度为55.07 ind./m2。综上所述,大型底栖动物在春、夏、秋3个季节的丰度低值区主要分布在中潮区和低潮区,最高值主要分布在高潮区。
从图3-B可见:大型底栖动物春季的生物量为3.58~23.94 g/m2,其中,C1站位生物量最高,D2站位最低,平均为13.77 g/m2;大型底栖动物夏季的生物量为8.89~48.76 g/m2,其中,A1站位最高,D2站位最低,平均生物量为24.55 g/m2;大型底栖动物秋季的生物量为7.00~32.37 g/m2,其中,A2站位最高,D2站位最低,平均为18.17 g/m2。
莱州湾南部胶莱河入海口处潮间带的大型底栖动物生物量与丰度的变化趋势总体一致,从季节上看,夏季生物量高于秋季和春季;从空间变化看,低潮区和中潮区大型底栖动物生物量在春、夏、秋3个季节均低于高潮区。
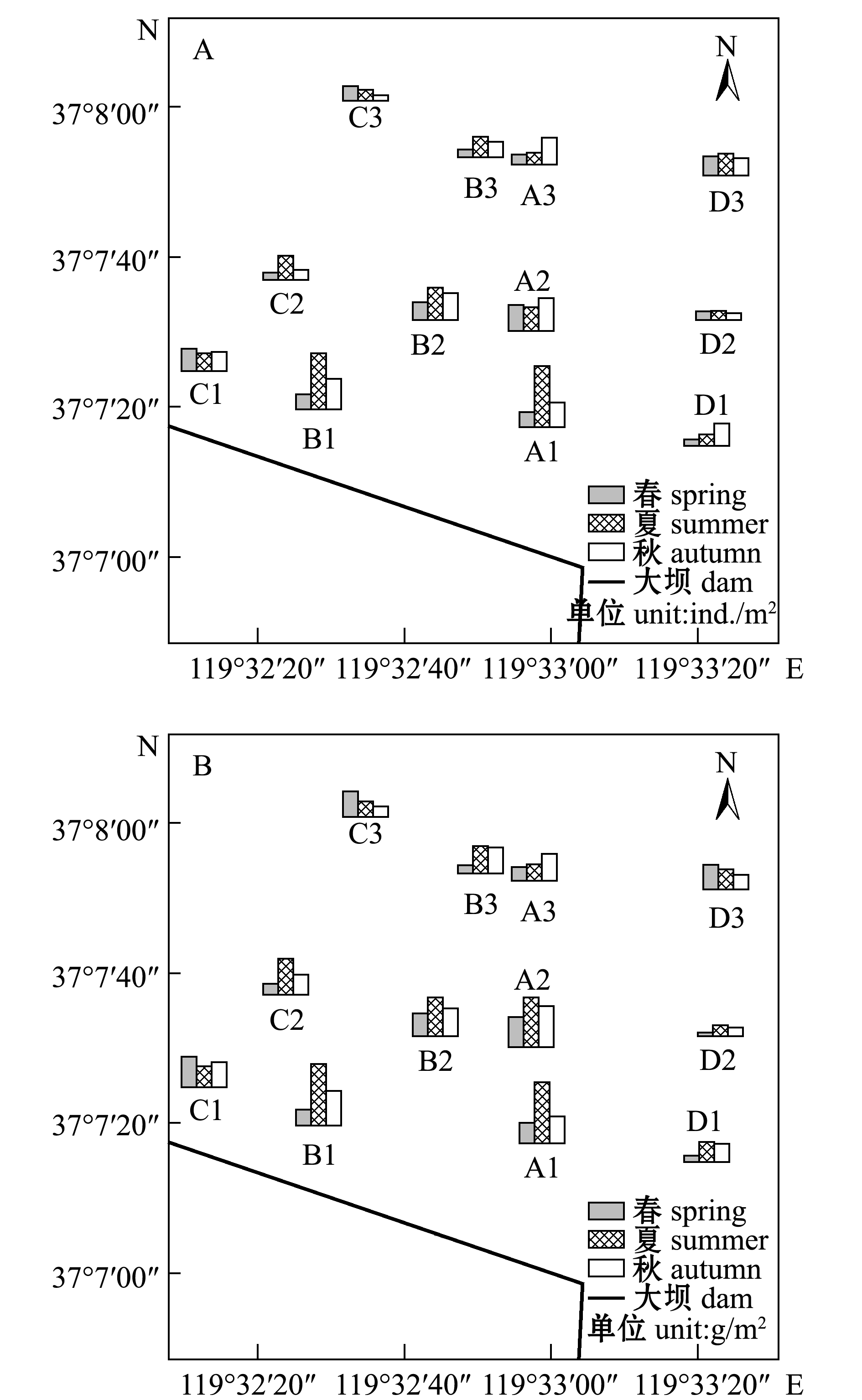
图3 不同季节大型底栖动物丰度(A)和生物量(B)的对比
Fig.3 Seasonal abundance (A) and biomass (B) of macrozoobenthos
表2显示了各个潮区大型底栖动物在不同季节的多样性指数。研究区内物种丰富度指数D平均为(1.25±0.06),变化范围为1.17~1.35,多样性指数H′平均为(2.33±0.15),变化范围为2.13~2.55,均匀度指数J平均为(0.67±0.05),变化范围为0.62~0.76。高潮区和低潮区的多样性指数H′在春季和秋季均高于中潮区,高潮区和低潮区的物种丰富度指数D平均值表现为春季和夏季高于秋季;各个潮区的多样性指数H′和均匀度指数J均表现为秋季和春季高于夏季,3个多样性指数在春季均表现为高潮区和低潮区大于中潮区。总体来看,莱州湾南部胶莱河入海口处潮间带大型底栖动物群落丰富度和多样性相对较好,而均匀度较低。
表2不同季节大型底栖动物多样性指数
Tab.2Differentseasondiversitiyindicesofmacrozoobenthos
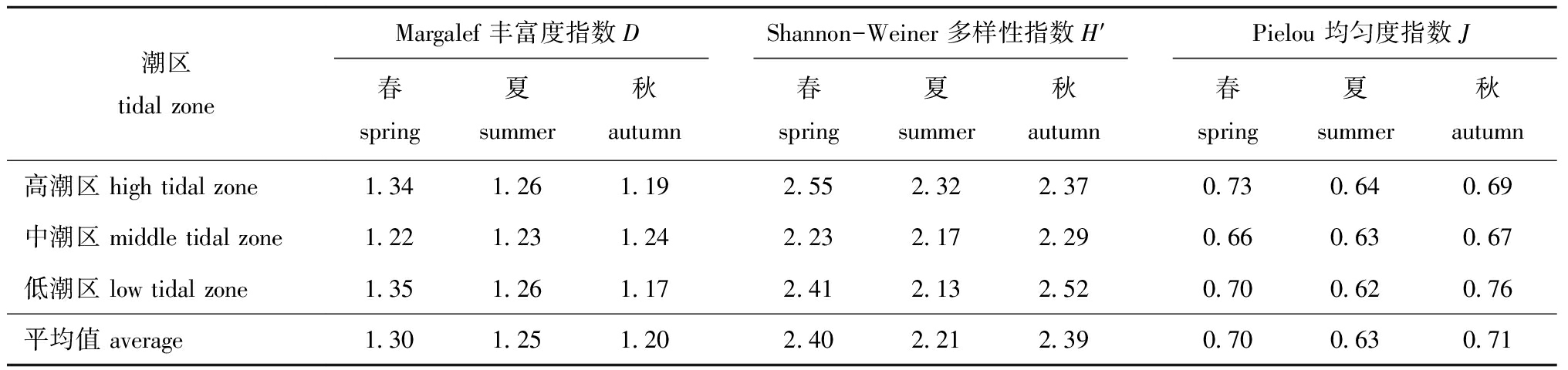
潮区tidal zoneMargalef 丰富度指数DShannon-Weiner 多样性指数H′Pielou 均匀度指数J春spring夏summer秋autumn春spring夏summer秋autumn春spring夏summer秋autumn高潮区 high tidal zone1.341.261.192.552.322.370.730.640.69中潮区 middle tidal zone1.221.231.242.232.172.290.660.630.67低潮区 low tidal zone1.351.261.172.412.132.520.700.620.76平均值 average1.301.251.202.402.212.390.700.630.71
软体动物为全年优势门类,占总丰度的90.52%,腕足动物和环节动物分别占4.57%和3.98%,节肢动物和原生动物丰度占比较少。从季节上看(图4),夏季软体动物的丰度比例最大,为92.64%,腕足动物占4.77%,环节动物占2.03%,节肢动物和原生动物分别占0.53%和0.04%;秋季软体动物丰度比例下降,为91.58%,腕足动物占4.06%,环节动物和节肢动物的比例上升,分别占比3.58%和0.76%,原生动物占0.03%;春季软体动物丰度所占比例最小,为85.29%,环节动物占8.0%,腕足动物和节肢动物分别占比4.93%和1.75%,原生动物占0.04%。
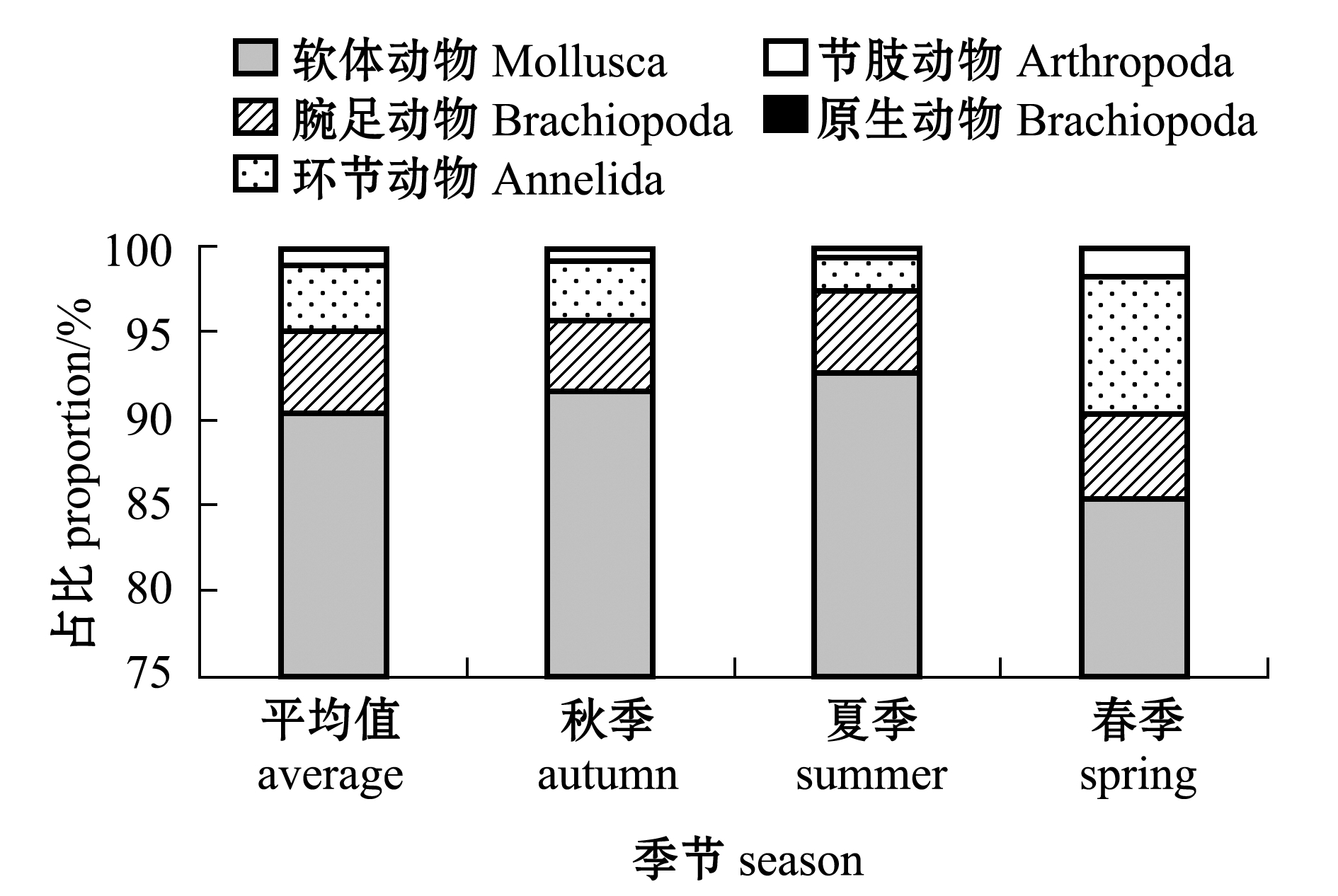
图4 各类群组成及季节变化
Fig.4 Species composition and variation of macrobenthos in different seasons
莱州湾南部胶莱河入海口处春季大型底栖动物优势种依次为异白樱蛤Macomaincongrua、红明樱蛤Moerellarutila、托氏虫昌螺Umboniumthomas;夏季和秋季大型底栖动物优势种基本相同,依次为异白樱蛤、红明樱蛤、托氏虫昌螺、古氏滩栖螺Batillariacumingi(表3)。3个常见种分别为大竹蛏Solengrandis、秀丽织纹螺Nassariusfestivus和青蛤Cyclinasinensis。
表3不同季节大型底栖动物优势种和常见种及优势度
Tab.3Dominantandcommonspeciesandtheirdominancesofmacrozoobenthosindifferentseasons
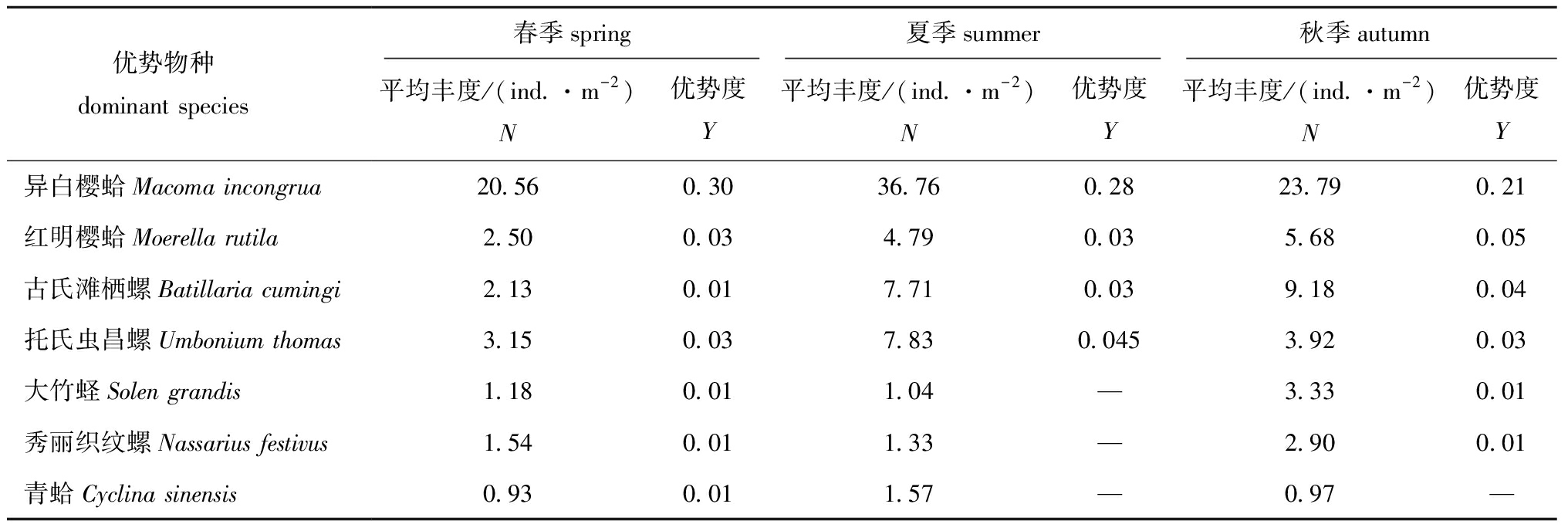
优势物种 dominant species 春季spring夏季summer秋季autumn平均丰度/(ind.·m-2)N优势度Y平均丰度/(ind.·m-2)N优势度Y平均丰度/(ind.·m-2)N优势度Y异白樱蛤Macoma incongrua20.560.3036.760.2823.790.21红明樱蛤Moerella rutila2.500.034.790.035.680.05古氏滩栖螺Batillaria cumingi2.130.017.710.039.180.04托氏虫昌螺Umbonium thomas3.150.037.830.0453.920.03大竹蛏Solen grandis1.180.011.04—3.330.01秀丽织纹螺Nassarius festivus1.540.011.33—2.900.01青蛤Cyclina sinensis0.930.011.57—0.97—
异白樱蛤,春季丰度为6.00~44.67 ind./m2,平均丰度为20.56 ind./m2;夏季为9.33~108.00 ind./m2,平均丰度为36.76 ind./m2;秋季为5.67~50.00 ind./m2,平均丰度为23.79 ind./m2;异白樱蛤为春、夏和秋3个季节共有的第一优势种,分别占大型底栖动物丰度的48.17%、48.89%和39.87%。
红明樱蛤,春季丰度为0.50~4.50 ind./m2,平均丰度为2.50 ind./m2;夏季为1.33~13.00 ind./m2,平均丰度为4.79 ind./m2;秋季为0.50~11.50 ind./m2,平均丰度为5.68 ind./m2,占大型底栖动物丰度的5.46%,为秋季第二优势种。
托氏虫昌螺,春季丰度为0~9.00 ind./m2,平均丰度为3.15 ind./m2,占大型底栖动物丰度的7.38%,为春季第二优势种;夏季为0~30.50 ind./m2,平均丰度为7.83 ind./m2,占大型底栖动物丰度的10.41%,为夏季第二优势种;秋季为0.17~10.00 ind./m2,平均丰度为3.92 ind./m2。
古氏滩栖螺,春季丰度为0~9.50 ind./m2,平均丰度为2.13 ind./m2,占大型底栖动物丰度的5.86%,在春季未能形成优势种;夏季为0~30.17 ind./m2,平均丰度为7.71 ind./m2;秋季为0~50 ind./m2,平均丰度为9.18 ind./m2,占大型底栖动物丰度的18.46%,为秋季第三优势种。
大竹蛏,春季丰度为0~4.83 ind./m2,平均丰度为1.18 ind./m2;夏季为0~4.5 ind./m2,平均丰度为1.04 ind./m2;秋季为0~14.00 ind./m2,平均丰度为3.33 ind./m2;春季、夏季和秋季分别占大型底栖动物总丰度的3.32%、1.67%和6.70%。
秀丽织纹螺,春季丰度为0~1.68 ind./m2,平均丰度为1.54 ind./m2;夏季为0.17~1.45 ind./m2,平均丰度为1.33 ind./m2;秋季为0~3.17 ind./m2,平均丰度为2.90 ind./m2;春季、夏季和秋季分别占大型底栖动物丰度的3.94%、1.93%和5.30%。
青蛤,春季丰度为0~3.17 ind./m2,平均丰度为0.93 ind./m2;夏季为0~8.00 ind./m2,平均丰度为1.57 ind./m2;秋季为0~3.83 ind./m2,平均丰度为0.97 ind./m2;春季、夏季和秋季分别占大型底栖动物丰度的2.38%、2.28%和1.78%。
ANOSIM相似性分析结果如表4所示,莱州湾南部胶莱河入海口处大型底栖动物群落结构在潮区之间存在着极显著性差异(P<0.01)。群落聚类分析结果如图5所示,在70%的相似性水平上可以分为3组:第一组,中潮区SPD2和SUD2聚为一支;第二组,高潮区(SPA1、SPB1、SPC1、SPD1、SUA1、SUB1、SUC1、SUD1、AUA1、AUC1、AUD1)和低潮区(SUA3)聚为一支;第三组,低潮区(SUD3、AUD3、SPA3、SPB3、SPD3、AUB3、SPC3、SUC3、AUA3、AUC3、SUB3)、中潮区(AUD2、AUC2、SPC2、AUA2、SUC2、SPA2、SUA2、AUB2、SPB2、SUB2)和高潮区的AUB1聚为一支。NMDS多维排序结果显示,其胁迫系数stress为0.16,说明MDS图可以解释底栖动物群落结构,进一步支持了聚类分析结果。ANOSIM分析表明,潮区之间的群落有极显著性差异(P<0.01)。
表4各潮区相似性矩阵相关系数
Tab.4Correlationcoefficientofsimilaritymatrixatdifferenttidalzones
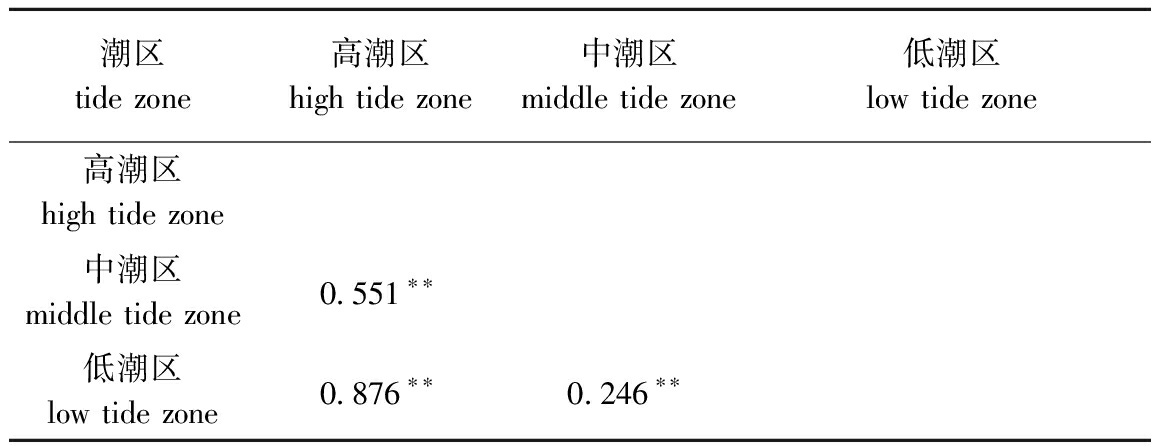
潮区 tide zone高潮区 high tide zone中潮区 middle tide zone低潮区 low tide zone高潮区 high tide zone中潮区 middle tide zone0.551∗∗低潮区 low tide zone0.876∗∗0.246∗∗
注: **表示极显著相关(P<0.01)
Note:**means very significant correlations(P<0.01)
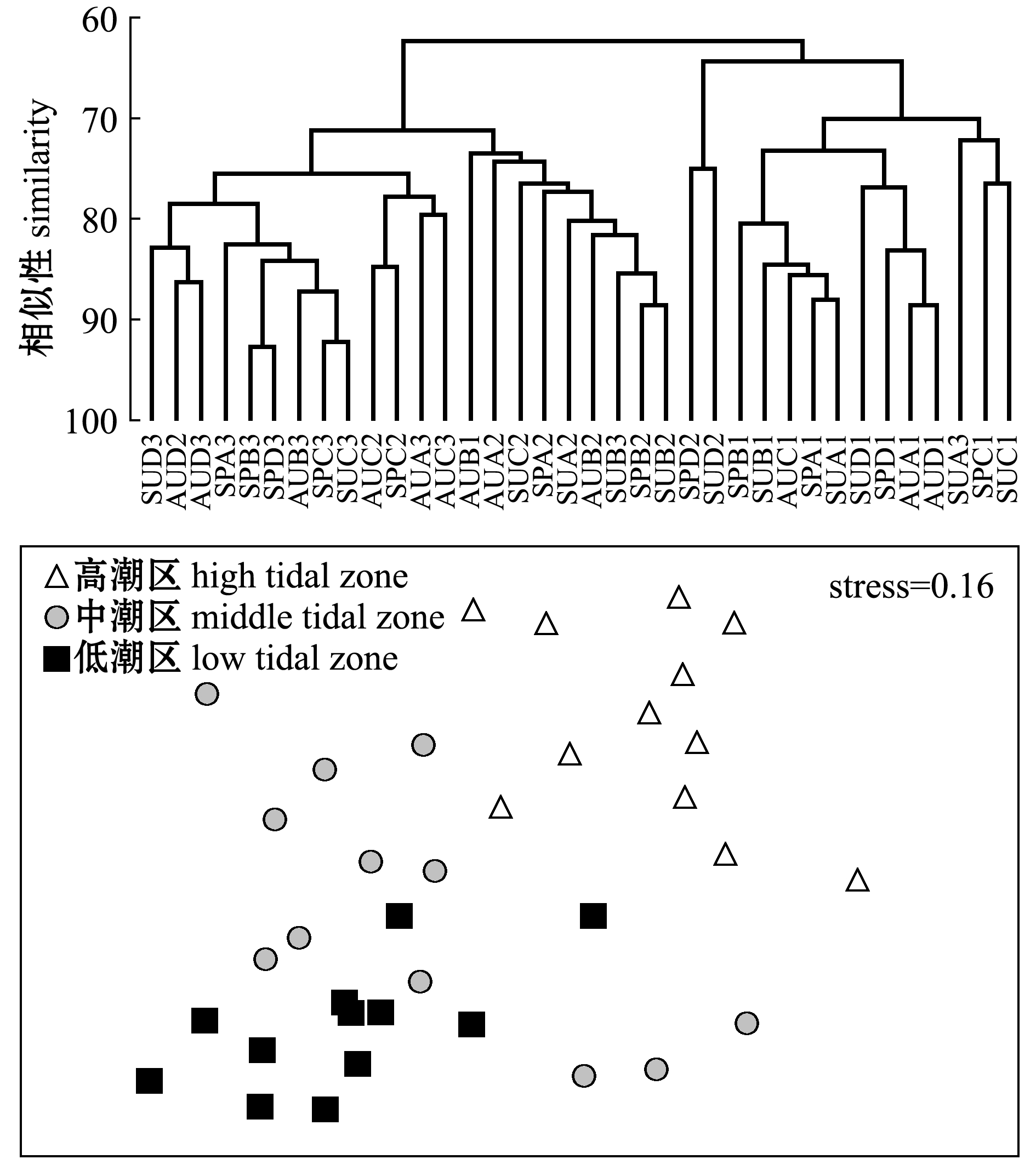
注:前两个字母代表季节,第三个字母代表站位,后面数字代表潮区;SP代表春季,SU代表夏季,AU代表秋季,1代表高潮区,2代表中潮区,3代表低潮区
Notes:the front 2 letters mean season,the third letter means site,and the last number means tidal zone;SP means spring,SU means summer,AU means autumn,1 means high tide zone,2 means middle tide zone,3 means low tide zone
图5 群落聚类分析和非度量多维度排序分析
Fig.5 Cluster analysis and NMDS of macrozoobenthos community
SIMPER相似性百分比分析结果显示(表5):莱州湾南部胶莱河口潮间带高潮区大型底栖动物组内平均相似性为74.31%,共有9种相似性累积贡献率超过90%,主要有异白樱蛤、古氏滩栖螺和红明樱蛤等,其中异白樱蛤的相似性累积贡献率最高;中潮区大型底栖动物组内平均相似性为70.47%,共有9种相似性累积贡献率超过90%,主要有异白樱蛤、红明樱蛤和海豆芽等,异白樱蛤的相似性贡献率仍最高;低潮区大型底栖动物组内平均相似性为77.96%,共有7种相似性累积贡献率超过90%,主要有异白樱蛤、托氏虫昌螺和红明樱蛤等,与高潮区和中潮区一样,异白樱蛤相似性贡献率均为最高。
各潮区大型底栖动物组成组间相异性结果显示(表6):高、中潮区组间平均相异性为35.57%,其中分歧种古氏滩栖螺、托氏虫昌螺、大竹蛏和薄壳绿螂的相异性累积贡献率达到35.31%;高潮区与低潮区组间平均相异性为39.84%,分歧种古氏滩栖螺、大竹蛏、薄壳绿螂的相异性累积贡献率达到32.65%;中潮区与低潮区组间平均相异性为28.78%,分歧种古氏滩栖螺、大竹蛏、托氏虫昌螺的相异性累积贡献率达到28.82%。综上所述,高潮区、中潮区和低潮区大型底栖动物的群落结构有明显差异。
表5各潮区大型底栖动物典型种及其对组内相似性的贡献率(>4%)
Tab.5Typicalspeciesandcontributionratesofintragroupsimilarityinmacrozoobenthoscommunityinvarioustidezones
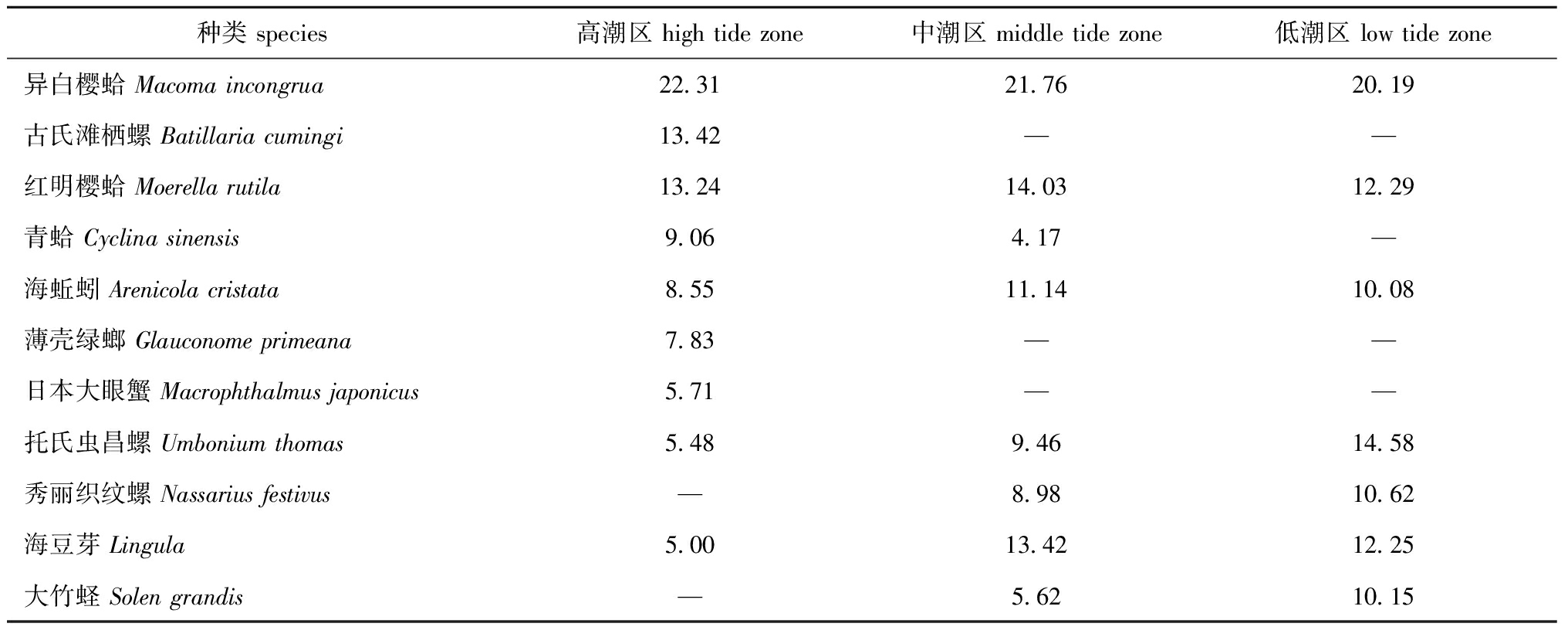
种类 species高潮区 high tide zone中潮区 middle tide zone低潮区 low tide zone异白樱蛤 Macoma incongrua22.3121.7620.19古氏滩栖螺 Batillaria cumingi13.42——红明樱蛤 Moerella rutila13.2414.0312.29青蛤 Cyclina sinensis9.064.17—海蚯蚓 Arenicola cristata 8.5511.1410.08薄壳绿螂 Glauconome primeana7.83——日本大眼蟹 Macrophthalmus japonicus5.71——托氏虫昌螺 Umbonium thomas5.489.4614.58秀丽织纹螺 Nassarius festivus—8.9810.62海豆芽 Lingula5.0013.4212.25大竹蛏 Solen grandis—5.6210.15
表6各潮区大型底栖动物分歧种及其对组间相异性的贡献率(>4%)
Tab.6Discriminatingspeciesandcontributionratesofintragroupsimilarityinmacrozoobenthoscommunityinvarioustidezones
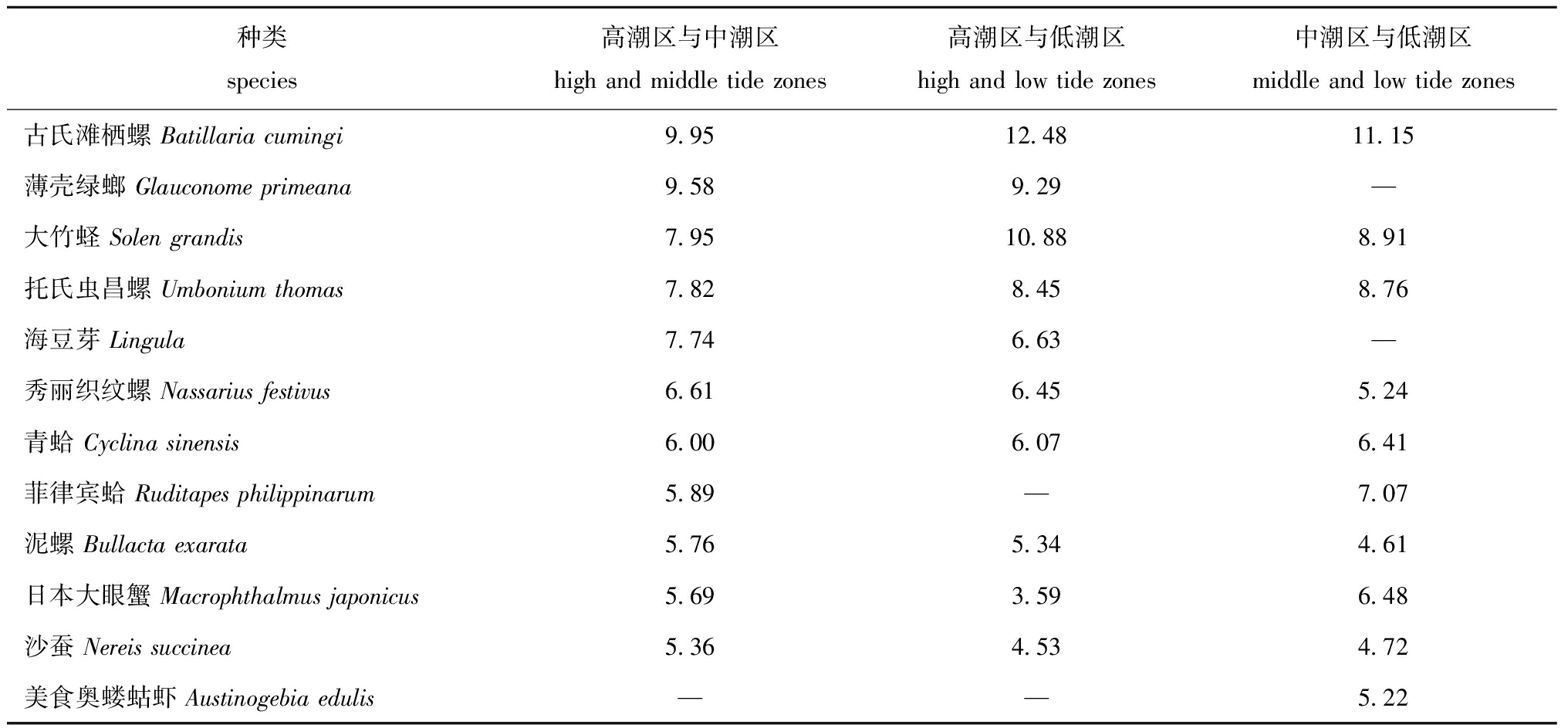
种类species高潮区与中潮区high and middle tide zones高潮区与低潮区high and low tide zones中潮区与低潮区middle and low tide zones古氏滩栖螺 Batillaria cumingi9.9512.4811.15薄壳绿螂 Glauconome primeana9.589.29—大竹蛏 Solen grandis7.9510.888.91托氏虫昌螺 Umbonium thomas7.828.458.76海豆芽 Lingula7.746.63—秀丽织纹螺 Nassarius festivus6.616.455.24青蛤 Cyclina sinensis6.006.076.41菲律宾蛤 Ruditapes philippinarum5.89—7.07泥螺 Bullacta exarata5.765.344.61日本大眼蟹 Macrophthalmus japonicus5.693.596.48沙蚕 Nereis succinea5.364.534.72美食奥蝼蛄虾 Austinogebia edulis——5.22
研究区域潮间带大型底栖生物群落结构的季节间存在一定的变化。从本次调查结果分析,各季节获得的物种数相对稳定(春季23种,夏季和秋季各24种),但是从生物量和丰度来看,夏季最高,秋季次之,春季最低,这表明季节对研究区潮间带大型底栖生物的生物量和丰度有一定的影响。林颂[15]研究表明,水温是影响大型底栖动物群落结构特征的重要因素之一,会引起沉积物环境特征、海流、溶解氧浓度等一系列变化,在大型底栖动物适宜的温度范围内,其生物量会随着水温的升高而增加[16-17]。春季繁殖个体较小且轻(特别是异白樱蛤、红明樱蛤、古氏滩栖螺等3个季节共有的优势种),夏季海水表层温度升高,营养盐丰富,微生物、水藻比较繁盛,给大型底栖动物提供了丰富的食物来源,此时底栖动物生长也较快,增重明显。Giliarov[18]认为,人类对潮间带经济种类的大量采集和挖掘底质可能是秋季潮间带生物量低的主要原因,秋季底栖生物由于被捕食等原因导致的死亡,使秋季的丰度和生物量均低于夏季。这与冷宇等[19]得出的秋季生物量较高的研究结果不同,原因可能是虽然研究区均位于莱州湾南部胶莱河口周围,但冷宇等的研究区在昌邑海洋生态特别保护区内部,受人为经济采捕干扰较小,故秋季生物量较大。
本研究中发现,大型底栖动物的丰度和生物量在潮区间存在差异,表现为高潮区大于中潮区和低潮区。袁兴中[20]研究发现,大型底栖动物物种数目和生物量明显受潮汐和波浪的作用影响,研究区内的优势种多为行动能力较弱且个体较小的软体动物(如异白樱蛤、红明樱蛤、海豆芽、大竹蛏等),所以与其他环境因子相比,波浪作用对大型底栖动物在潮间带分布的影响也较大[21]。由于受到人类采捕需要挖坑等活动影响,高潮区的地形变化和微地貌元素比中潮区和低潮区更为复杂,它们对底栖动物的丰度格局和多样性有着重要的影响[20],部分大型底栖动物由于水动力作用经泥沙裹挟被冲刷至高潮区,但在落潮回流时因高潮区内凹凸不平的地形滞留了部分大型底栖动物,且这些物种大多属于可以忍受阳光照射及每日二次涨退潮干湿变化的物种[22],所以这些来自低潮区和中潮区的大型底栖生物也可以在高潮区繁衍生存。张志南等[23]、李新正等[24]指出,混合沉积环境的空间异质化程度更高,可以为底栖动物的生长和发育提供更多的空间,这也是生物多样性较高的基础。经实地调查,潮水搬运泥沙在高潮区沉积后,形成南高北低(即高潮区高低潮区低)的地势格局,大量泥沙堆积致使高潮区相对于低潮区沉积环境更为复杂,且高潮区大量泥沙形成的混合质沉积物,为埋栖生活的小型双壳类、甲壳类、浅穴居和小型多毛类动物提供了丰富的食物和良好的栖息环境,使得大型底栖动物在此大量繁衍生存[2]。
基于Bray-Curtis相似性系数的ANOSIM检验表明,莱州湾胶莱河口潮间带大型底栖动物群落结构在潮区之间存在显著性差异。群落结构的差异主要体现在群落典型种差异性,SIMPER分析表明,除了异白樱蛤和红明樱蛤为3个潮带共有的典型种外,高潮区的典型种主要是古氏滩栖螺、青蛤、日本大眼蟹和薄壳绿螂,中潮区主要是海豆芽、海蚯蚓和托氏虫昌螺,低潮区主要是托氏虫昌螺、海豆芽、秀丽织纹螺、大竹蛏和海蚯蚓,其中海蚯蚓为污染指示种。多数组内典型种又是组间分歧种,是不同潮区群落资源量占优势的种类。
蔡立哲等[25]使用多样性指数H′值分级评价标准进行环境污染评价,当1<H′<2时表示中度污染,2<H′<3时表示轻度污染,H′>3时表示环境清洁。本研究中,春、夏、秋3个季节的H′平均值为2~39,结合上述污染指示种海蚯蚓为低潮区和中潮区出现的共同典型种,表明研究区内环境已受到轻微污染。本次调查中秋季多样性指数H′的平均值(2.39)高于高昊东等[6]的调查结果(1.33),原因可能是高昊东等研究得出的胶莱河口东岸潮间带间隙水中存在高溴等污染物的结果与东岸分布的盐化工厂和工业区有关[6],而本研究区所处的西岸潮间带所受污染相对较小。
2009年秋季(10月)高昊东等[6]在莱州湾南部胶莱河入海口西岸潮间带采样得到大型底栖动物平均丰度和平均生物量分别为380 ind./m2和57.40 g/m2,2011年秋季(9月)顾炎斌等[7]在东营市河口区潮间带取样得到大型底栖动物平均丰度和平均生物量分别为159.11 ind./m2和186.22 g/m2。本研究区内秋季大型底栖动物平均丰度和平均生物量分别为55.07 ind./m2和18.17g/m2,对比发现,本研究区内生物量水平相对较低,生境不稳定是生物量下降的重要原因[26],潮间带作为陆海交互的系统单元,其稳定性并不高,而且易受人类活动的影响[22],实地调查发现,人类对潮间带经济种类的开发、对底质的采掘和陆源排污可能是潮间带大型底栖动物丰度和生物量逐年降低的主要原因,加上研究区内多次实施围填海等大工程建设,使莱州湾胶莱河口潮间带大型底栖动物生态指数偏低。高昊东等[6]、顾炎斌等[7]和林颂[15]均发现,除了软体动物,滩涂底栖动物优势种还包括多毛类和甲壳类。本研究中,软体动物类成为全年内单一优势类群,且不同月份种类变化数也较小,可能是由于研究区内滩涂受人类采贝等活动干扰加剧,导致生命周期长和个体较大的甲壳类等减少,而生命周期短和个体较小的异白樱蛤、古氏滩栖螺等小型软体动物大量繁殖[16]。
潮汐和生境差异导致潮间带空间的异质性(沉积物组成差异),空间异质性导致大型底栖动物群落结构的差异。莱州湾南部胶莱河入海口处潮间带大型底栖动物受到沉积物差异、温度、盐度、地形等生境差异的影响,从而呈现出季节性差异和潮区间差异。由于潮间带生态环境复杂多变,且影响大型底栖动物的环境因子繁多,因此,较难用一种或者几种因素的变化来解释大型底栖动物的变化[26],而且大型底栖动物在环境发生变化的同时,自身也在不断适应、调整和变化,因此,对大型底栖动物的研究需要进行更长期、深入地调查研究以发现其规律。
参考文献:
[1] Desrosiers G,Savenkoff C,Olivier M,et al.Trophic structure of macrobenthos in the Gulf of St.Lawrence and on the Scotian Shelf[J].Deep-Sea Research Part II:Topical Studies in Oceanography,2000,47(3-4):663-697.
[2] Soko owski A,Woowicz M,Asmus H,et al.Is benthic food web structure related to diversity of marine macrobenthic communities?[J].Estuarine,Coastal and Shelf Science,2012,108:76-86.
owski A,Woowicz M,Asmus H,et al.Is benthic food web structure related to diversity of marine macrobenthic communities?[J].Estuarine,Coastal and Shelf Science,2012,108:76-86.
[3] Thorin S,Radureau A,Feunteun E,et al.Preliminary results on a high east-west gradient in the macrozoobenthic community structure of the macrotidal Mont Saint-Michel bay[J].Continental Shelf Research,2001,21(18-19):2167-2183.
[4] 赵永强,曾江宁,高爱根,等.椒江口滩涂大型底栖动物群落格局与多样性[J].生物多样性,2009,17(3):303-309.
[5] 王宝强,薛俊增,庄骅,等.洋山港潮间带大型底栖动物群落结构及多样性[J].生态学报,2011,31(20):5865-5874.
[6] 高昊东,罗新正,孙万龙,等.莱州湾胶莱河口潮间带大型底栖动物群落生态学研究[J].齐鲁渔业,2011(2):1-6.
[7] 顾炎斌,宋文豪,姚韡远,等.东营市河口区潮间带大型底栖动物群落结构特征[J].海洋环境科学,2013,32(5):702-706.
[8] 国家质量技术监督局发布.GB 17378.7-1998 海洋监测规范第7部分:近海污染生态调查和生物监测[M].北京:中国标准出版社,1998.
[9] 周红,张志南.大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J].中国海洋大学学报:自然科学版,2003,33(1):58-64.
[10] 徐兆礼,陈亚瞿.东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J].生态学杂志,1989,8(4):13-15.
[11] Khalaf M A,Kochzius M.Changes in trophic community structure of shore fishes at an industrial site in the Gulf of Aqaba,Red Sea[J].Marine Ecology Progress Series,2002,239(1):287-299.
[12] 斯广杰,陈丕茂,杜飞雁,等.深圳杨梅坑人工鱼礁区投礁前后大型底栖动物种类组成的变化[J].大连海洋大学学报,2010,25(3):243-247.
[13] 马藏允,刘海,王惠卿,等.底栖生物群落结构变化多元变量统计分析[J].中国环境科学,1997,17(4):297-300.
[14] 陈强,郭行磐,周轩,等.长江口及其邻近水域滩涂底栖动物多样性的研究[J].大连海洋大学学报,2016,31(1):103-108.
[15] 林颂.小清河口及邻近海域沉积环境特征及底栖生境质量评价[D].青岛:中国海洋大学,2015.
[16] Juario J V.Nematode species composition and seasonal fluctuation of a sublittoral meiofauna community in the German Bight[J].Veroff Inest Meeresforsch Bremerh,1975,11(2):153-159.
[17] 卓异,蔡立哲,郭涛,等.泉州湾蟳埔潮间带大型底栖动物群落的时空分布[J].生态学报,2014,34(5):1244-1252.
[18] Giliarov A M.Information theory in ecology[J].Computers & Chemistry,2001,25(4):393-399.
[19] 冷宇,张守本,刘一霆,等.昌邑海洋生态特别保护区潮间带大型底栖动物时空分布特征[J].中国环境监测,2014,30(2):164-169.
[20] 袁兴中.河口潮滩湿地底栖动物群落的生态学研究[D].上海:华东师范大学,2001.
[21] 杨万喜,陈永寿.嵊泗列岛潮间带群落生态学研究Ⅲ.岩相潮间带底栖生物的种类分布[J].海洋学研究,1999,17(1):60-65.
[22] 仇建标,彭欣,谢起浪,等.洞头列岛潮间带大型底栖生物的时空分布及其多样性[J].水产学报,2012,36(4):608-614.
[23] 张志南,林岿旋,周红,等.东、黄海春秋季小型底栖生物丰度和生物量研究[J].生态学报,2004,24(5):997-1005.
[24] 李新正,于海燕,王永强,等.胶州湾大型底栖动物的物种多样性现状[J].生物多样性,2001,9(1):80-84.
[25] 蔡立哲,马丽,高阳,等.海洋底栖动物多样性指数污染程度评价标准的分析[J].厦门大学学报:自然版,2002,41(5):641-646.
[26] 施华宏.滨海采捕对岸带生物资源的影响及管理对策[J].海洋开发与管理,2001,18(2):32-34.
LI Hui-feng1,2, YANG Wen-bo1,2, YUAN Li-lai1, LIU Bao-xiang1, CAO Kun1, WANG Lin1, DING Fang1, LI Hui-qin1, LI Ji-long1, LI Ying-ren1, HUANG Jing-xian3, ZHANG Chuan-tao3
(1.Fisheries Resource and Environment Research Center, Chinese Academy of Fishery Sciences, Beijing 100141, China; 2.College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China; 3.Xiaying Enhancement and Experiment Station, Chinese Academy of Fishery Sciences, Changyi 261300, China)
Abstract:The seasonal variation and community structure of intertidal macrozoobenthos was studied based on the data collected from Jiaolai estuary of south Laizhou Bay in spring (April of 2016),summer (July of 2016) and autumn(October of 2016). A total of 28 species belonging to 5 phyla were found, including Mollusca (19 species), Arthropoda (5 species), Annelida(2 species), Brachiopoda (2 species) and Protozoa(1 species). The average resources were shown to be 13.77 g/m2in biomass density and 38.89 ind./m2in number in spring, and 24.55 g/m2in biomass density and 68.49 ind./m2in number in summer, and 18.17 g/m2in biomass density and 55.07 ind./m2in number in autumn. Mollusca and Arthropoda were the dominant groups, with dominant speciesMacomaincongrua,Moerellarutila, andUmboniumthomasin spring, andMacomaincongrua,Moerellarutila,Umboniumthomas, andBatillariacumingias dominant species both in summer and autumn. The diversity index (H′)was ranged from 2.13 to 2.55, the Pielou evenness index (J)from 0.62 to 0.73, and the Margalef species richness index (D)from 1.17 to 1.35. ANOSIM analysis, cluster and NMDS analysis revealed that there was different macrozoobenthic community structure in tide zones in Jiaolai estuary of south Laizhou Bay. The SIMPER analysis indicated thatMacomaincongruaandMoerellarutilawere the typical species, andBatillariacumingi,Solengrandis, andGlauconomeprimeanawere the discriminating species.
Keywords:intertidal zone; macrozoobenthos; spatial and temporal distribution; community structure
通信作者:李继龙(1958—), 男, 研究员。E-mail:lijilong@cafs.ac.cn
作者简介:李慧峰(1992—), 女, 硕士研究生。E-mail:799313886@qq.com
基金项目:中国水产科学研究院基本科研业务费(2016B003);农业部物种资源保护费项目(2130135)
收稿日期:2017-12-01
文章编号:2095-1388(2018)05-0674-09
DOI:10.16535/j.cnki.dlhyxb.2018.05.020
文献标志码:A
中图分类号:Q178.1