血鹦鹉俗称红财神、财神鱼、发财鱼,该鱼最早来自中国台湾,由鲈形目Perciformes、慈鲷科Cichlidae的红头丽体鱼 Vieja synspila(Hubbs,1935)♀和橘色双冠丽体鱼Amphilophus citrinellus(Gunther,1864)♂杂交而来。其体色红艳,体型圆润,嘴呈心形,笑口常开,深受广大消费者的喜爱,已成为世界范围内重要的淡水经济观赏鱼类。中国血鹦鹉鱼的养殖区域主要集中于天津、上海、广东、海南等地。天津地区年产量可达10 940万尾,产值约为9788万元,占天津市观赏鱼年产量的9.95%,占观赏鱼产值的13.98%[1],有效带动了周边地区饲料、渔药、水族和运输等行业的发展。血鹦鹉属于远缘杂交子代,遗传性状不稳定,遗传差异较大,养殖过程中要选择褪色完全、体型圆润的个体作为商品鱼,尖头及不褪色或褪色不完全的血鹦鹉会在养殖过程中被淘汰,所以血鹦鹉尖头率的高低会直接影响企业及养殖户的经济效益。
在现有血鹦鹉繁育过程中,养殖户往往只通过鱼体外形挑选亲鱼,缺乏对亲本的种质评估环节。加上亲本的重复利用,使得繁育的血鹦鹉质量不稳定,经常出现尖头率高、褪色率低的现象,给养殖户造成了极大的经济损失。目前,关于血鹦鹉的研究主要集中于水质调控、养殖技术、饲料添加剂等方面[2-5],对血鹦鹉亲本及家系的遗传评估尚且较少,仅见刘肖莲等[6]利用微卫星标记对血鹦鹉3个全同胞家系进行遗传多样性及经济性状连锁分析。因此,亟需开展血鹦鹉及其父母本的种质遗传评估,以期为血鹦鹉分子育种提供参考。
微卫星标记具有多态性高、遗传稳定且呈共显性遗传等特性,该技术目前已广泛用于遗传多样性分析、遗传图谱的构建和分子育种等研究中[7-12]。本研究中,利用6对微卫星标记对2种体型的血鹦鹉家系进行种质遗传评估和分析,旨在为血鹦鹉亲本组配及分子育种提供理论依据。
1 材料与方法
1.1 材料
试验用紫红火口与红魔鬼鱼取自天津嘉禾田源观赏鱼养殖有限公司。
1.2 方法
1.2.1 试验设计 挑选性腺发育良好、体色鲜艳、健康无损伤的紫红火口 (♀)与红魔鬼 (♂)进行配对,自然产卵,待其产卵完成后将附着受精卵的瓦片放入孵化缸 (40 cm×30 cm×30 cm)中,微流水条件下孵化 (30.0±0.5)℃。当血鹦鹉苗种长至1.5 cm时转入苗种培育缸 (65 cm×40 cm×40 cm)中培育,投喂桡足类、枝角类或配合饲料(粗蛋白质含量为52%)。培育期间定期换水,以保持水质清新,pH为7.5~8.1,溶解氧保持在5.0 mg/L以上,水温为28~32℃,光照强度为1000 Lx。当血鹦鹉生长至3~4 cm时体色开始转色,体色由黑色渐变为淡橘色或浅黄色。当体长达到5 cm时,根据头型、体型、体色和嘴型进行苗种挑选,抽样统计尖头和圆头比率。选取完全褪色且尖头率低的家系 (YT)和尖头率高的家系 (JT)作为试验对象,每个家系随机选取30尾个体,测量其全长、体长、体高并计算体长/体高和全长/体高。剪取鳍条晾干后保存备用。
1.2.2 DNA提取及引物合成 本试验中所用微卫星标记参照刘肖莲等[6]开发的6个微卫星标记,标记名称、引物序列、退火温度和产物分布范围如表1所示。引物序列由上海生工生物工程技术服务有限公司合成。利用血液DNA提取试剂盒 (德国,QIAGEN)提取基因组DNA。提取完成后采用10 g/L琼脂糖凝胶电泳检测DNA样品的质量,于冰箱 (-20℃)中保存备用。
1.2.3 PCR扩增及基因分型 PCR反应体系 (共20 μL): 10×PCR buffer(Mg2+)2 μL, dNTP Mixture 1.6 μL, 正、 反向引物 (10 μmol/L) 各 1 μL, 5 U/μL Taq 酶0.5 μL, 50 ng/μL 基因组 DNA 1 μL,用ddH2O补足体积至20 μL。PCR反应程序为:95℃下预变性5 min;95℃下变性30 s,退火30 s(最佳退火温度见表1),72℃下延伸30 s,共进行30个循环;最后在72℃下再延伸10 min。PCR扩增产物用ABI3730基因分析仪 (Applied Biosystems)进行毛细管凝胶电泳,以GS500LIZ为长度内参分析产物长度,用3730 Data collection和GeneMapper 4.0软件读取微卫星扩增产物基因片段数据,并根据大小确定各样本在微卫星位点的基因型。
表1 微卫星引物序列及扩增情况
Tab.1 Sequence and amplification of microsatellite loci

位点locus引物序列primer退火温度/℃annealing temperature等位基因大小/bp size of allele Vsac1F:TGCCTCTCTTTCGGGTTT R:TTCCAGGATGGGATTGCT 50 174~186 Vsac2F:CGGGTTGTTCTGGACTTT R:AGATGTTGAGGTGGGTGC 58 359~416 Vsac3F:GATAGTCAAATGAGTTCAGCG R:CAGTAATCCGAAATCAAACG 58 224~233 Vsac4F:GCCAATCCTACAGAAAGATGAAA R:ACAATAAGCCACAAGCAATAAAAG 55 240~245 Vsac5F:TGGTAGGTTTGGTAATAGGA R:TTGCCAGTGGTTTCACAT 58 337~391 Vsac6F:CCCCATTGCTCTTCTGTA R:TTATCCGTCTCACCCTCA 53 226
1.3 数据处理
用PopGene 3.2软件统计微卫星位点的等位基因数 (Na)、有效等位基因数 (Ne)、观测杂合度(Ho)、期望杂合度 (He)、香农信息指数 (I)和固定系数 (Fis),使用PIC CALC 0.6软件计算各个位点的多态信息含量 (PIC),并检测各参数的平均值。使用SPSS 21.0软件对两种体型的血鹦鹉家系进行性状比值的正态分布检验 (Kolmogorov-Smimov法)和判别分析。利用广义线性模型(GLM)进行微卫星标记的最小二乘分析,筛选与经济性状显著相关的标记,建立模型如下:
其中:y为性状;u为群体平均值;g为第i个基因型的效应;e为残差。利用Duncan多重比较法分析与经济性状显著相关的微卫星位点不同基因型间的差异,并基于Permutation检验,确认基因型效应的显著性水平 (10 000次)。
2 结果与分析
2.1 血鹦鹉家系性状正态分布
通过紫红火口 (♀)与红魔鬼 (♂)进行配组共构建9个血鹦鹉全同胞家系。当血鹦鹉体长达到4~5 cm时,进行一次挑选,统计尖头情况,根据统计结果从中选取尖头 (尖头率84%)和圆头(尖头率10%)两个家系进行后续分析。
血鹦鹉家系体长/体高和全长/体高呈现连续变异的特点,采用SPSS 21.0软件中的单样本Kolmogorov-Smimov检验方法,检验两个家系性状比值的频率是否符合正态分布,结果见表2。经检验,YT家系的体长/体高、全长/体高及JT家系的体长/体高符合正态分布 (P>0.05),而JT家系的全长/体高不符合正态分布 (P<0.05)。从两个家系的体长/体高的分布频率 (表3)可知,这两个家系的血鹦鹉体型差异较大,YT家系的30尾个体的体长/体高为1.1~1.5,JT家系中93.3% 的个体体长/体高均大于1.5。两个血鹦鹉家系的性状比值分布见图1。
表2 血鹦鹉家系的性状比值的正态分布检验
Tab.2 Gaussian distribution of measurable traits of blood parrot fish families
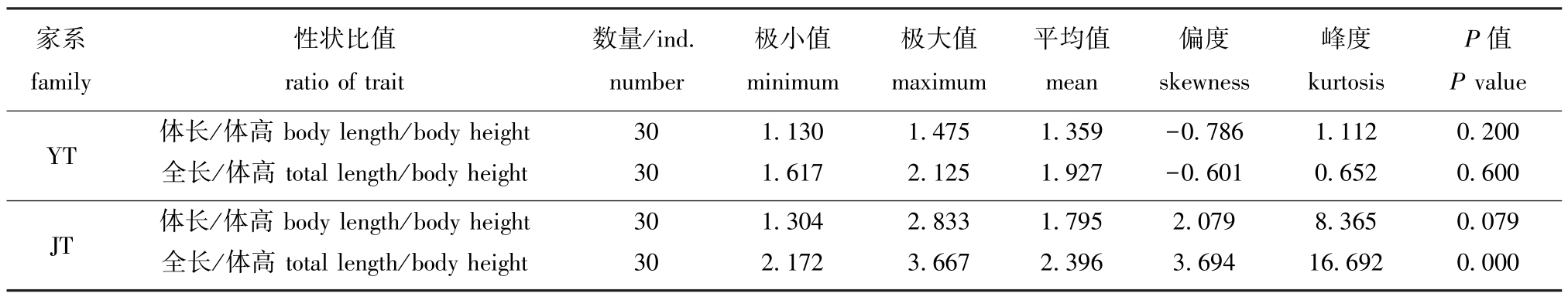
家系family性状比值ratio of trait数量/ind.number极小值minimum极大值maximum平均值mean偏度skewness峰度kurtosis P值P value YT 体长/体高 body length/body height 30 1.130 1.475 1.359 -0.786 1.112 0.200全长/体高 total length/body height 30 1.617 2.125 1.927 -0.601 0.652 0.600 JT 体长/体高 body length/body height 30 1.304 2.833 1.795 2.079 8.365 0.079全长/体高 total length/body height 30 2.172 3.667 2.396 3.694 16.692 0.000
表3 血鹦鹉家系个体体型分布情况
Tab.3 Distribution of body type of blood parrot fish families ind.

YT 0 0 1 6 16 7 0 30 JT 0 0 0 0 1 1 28 30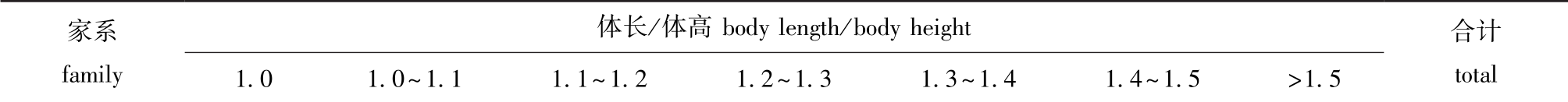
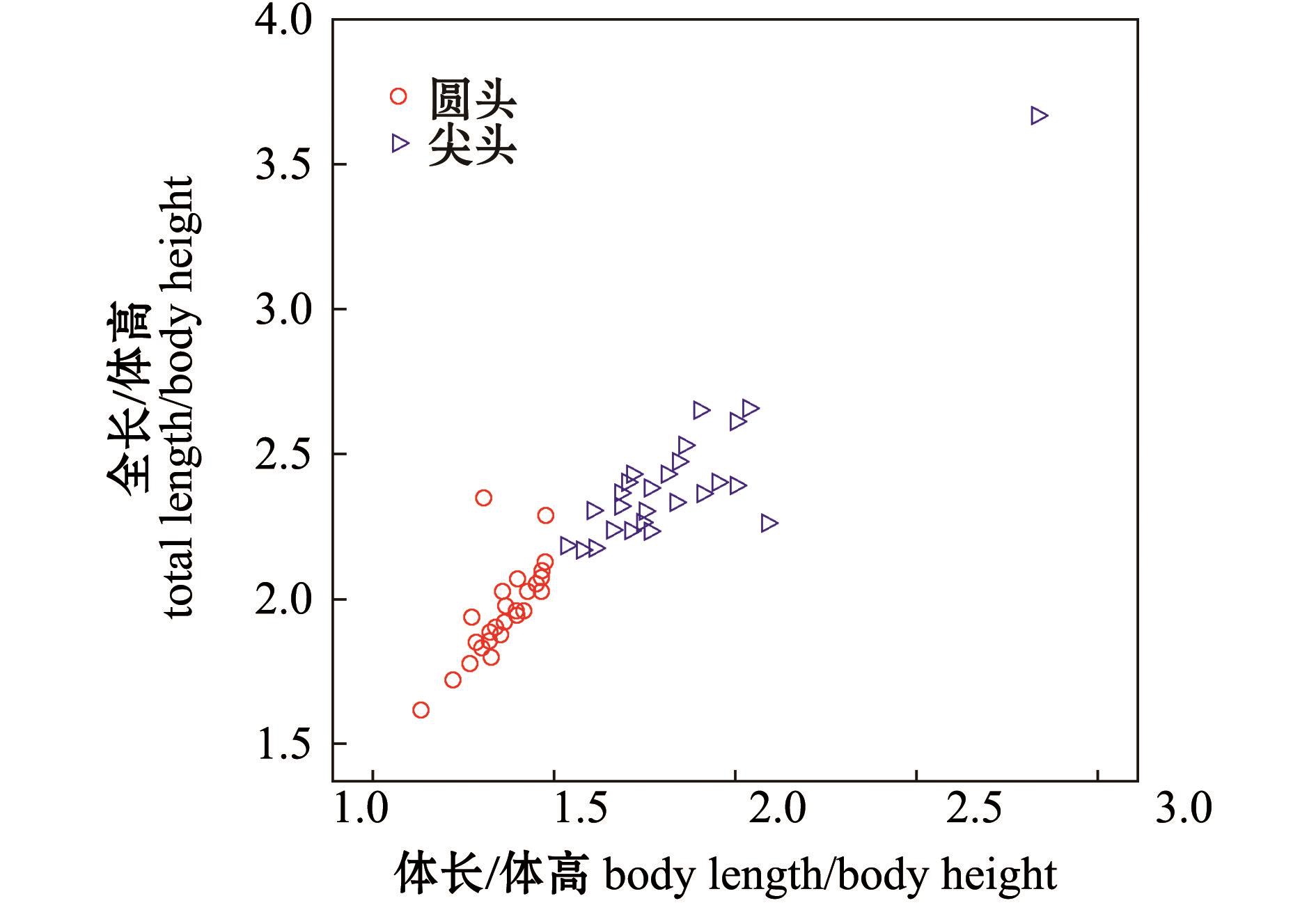
图1 两个血鹦鹉家系的性状比值分布
Fig.1 Scatter diagram of trait ratio in two blood parrot fish families
2.2 判别分析
按照体长/体高对血鹦鹉进行分组,比值大于1.50的定义为尖头 (28尾),比值小于1.5的定义为圆头 (32尾)。利用体长/体高(X1)和全长/体高(X2)的特征值构建判别公式为
在进行判别分析时,将所测2个特征值代入上述判别公式,所得函数值为正值即为尖头血鹦鹉,负值则为圆头血鹦鹉。据此对所有观测样本按照上述判别函数进行预测分类 (表4),YT家系血鹦鹉判别准确率为100%,JT家系判别准确率为89.3%,总体平均拟合概率为95%。因此,利用形态性状进行分类的判别分析成功率高,判别效果较好 (P<0.01)。
表4 血鹦鹉体型判别分析结果
Tab.4 Results of discriminant analysis of blood parrot fish
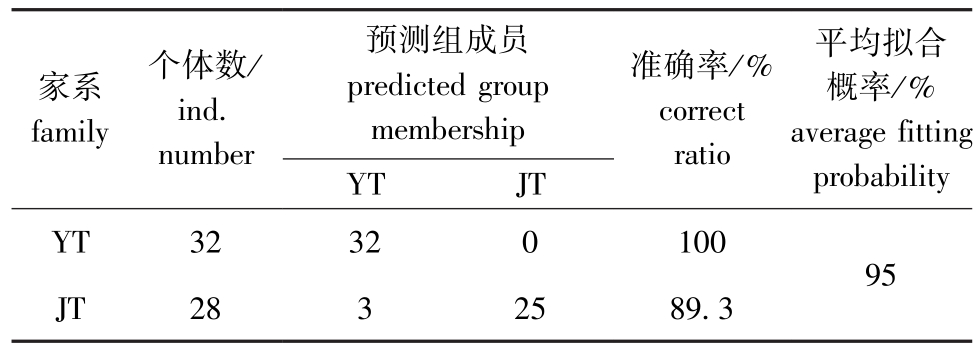
家系family个体数/ind.预测组成员predicted group membership准确率/%correct平均拟合概率/%average fitting YT 32 32 0 100 JT 28 3 25 89.3 95
2.3 血鹦鹉家系的遗传多样性
本研究中,所用6个微卫星标记均能在两个血鹦鹉家系中扩增出清晰稳定的DNA条带,扩增片段大小为174~416 bp。各个位点的等位基因数(Na)为 2~4个,有效等位基因数 (Ne)为1.032~3.908个,期望杂合度 (He)为 0.031~0.756,观测杂合度 (Ho)为0.031~0.969,多态信息含量 (PIC)为0.030~0.696(表5)。位点Vsac04和位点 Vsac06表现为低度多态 (PIC<0.25),位点Vsac05(YT家系)表现为高度多态(PIC>0.5),其余位点表现为中度多态 (0.25≤PIC≤0.5)。在固定系数 (Fis)方面,两个家系的Fis平均值分别为-0.488和-0.367,均表现为杂合子过剩 (Fis<0)。比较两个家系以上遗传参数发现, YT家系的 Ne、 He、 Ho、 PIC、 I和 Fis的平均值均高于JT家系,YT家系的遗传多样性较丰富。
表5 血鹦鹉家系在6个微卫星位点的遗传多样性
Tab.5 Genetic diversity of 6 microsatellite loci in blood parrot fish families
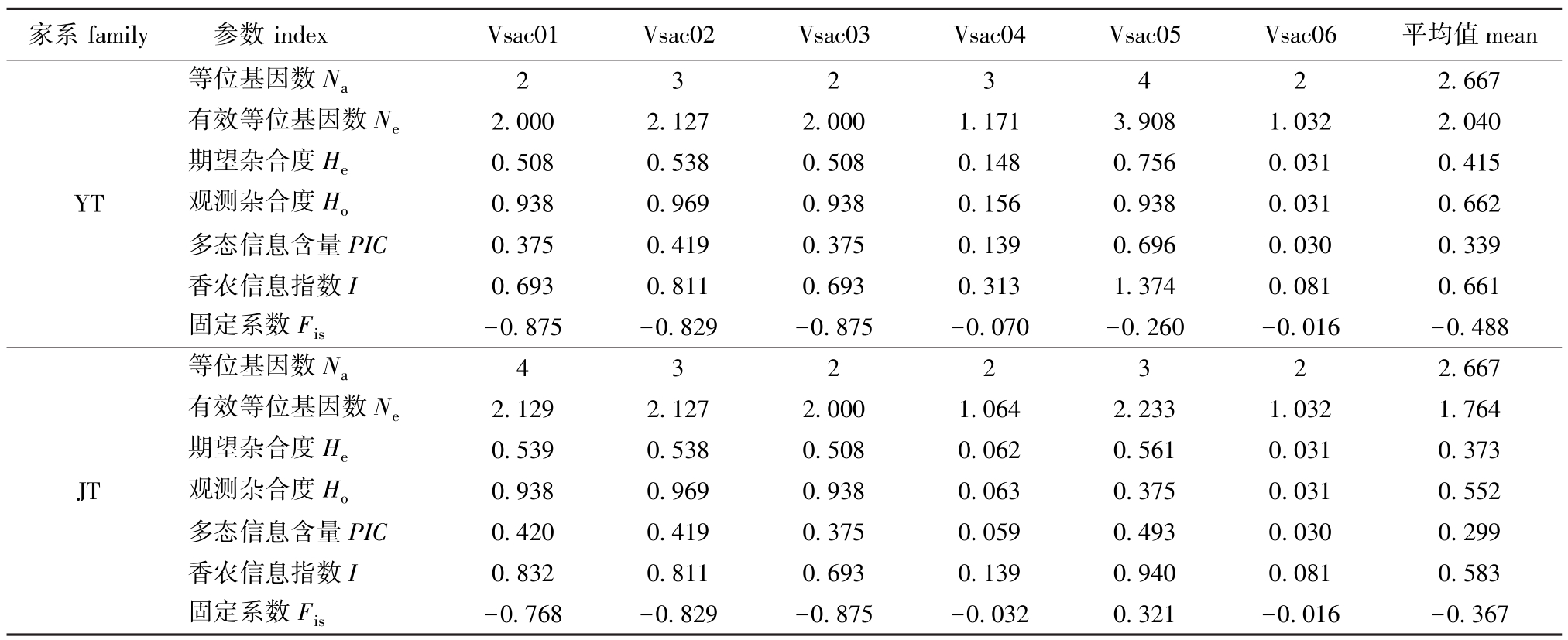
等位基因数Na 2 3 2 3 4 2 2.667有效等位基因数Ne 2.000 2.127 2.000 1.171 3.908 1.032 2.040期望杂合度He 0.508 0.538 0.508 0.148 0.756 0.031 0.415 YT 观测杂合度Ho 0.938 0.969 0.938 0.156 0.938 0.031 0.662多态信息含量PIC 0.375 0.419 0.375 0.139 0.696 0.030 0.339香农信息指数I 0.693 0.811 0.693 0.313 1.374 0.081 0.661固定系数Fis -0.875 -0.829 -0.875 -0.070 -0.260 -0.016 -0.488等位基因数Na 4 3 2 2 3 2 2.667有效等位基因数Ne 2.129 2.127 2.000 1.064 2.233 1.032 1.764期望杂合度He 0.539 0.538 0.508 0.062 0.561 0.031 0.373 JT 观测杂合度Ho 0.938 0.969 0.938 0.063 0.375 0.031 0.552多态信息含量PIC 0.420 0.419 0.375 0.059 0.493 0.030 0.299香农信息指数I 0.832 0.811 0.693 0.139 0.940 0.081 0.583固定系数Fis -0.768 -0.829 -0.875 -0.032 0.321 -0.016 -0.367
2.4 微卫星标记与性状的关联分析
采用SPSS 21.0软件计算的GLM模型,对微卫星标记和体长/体高、全长/体高性状进行相关性分析,结果表明,标记Vsac04和Vsac05与这两个性状显著相关 (P≤0.05),但Vsac04只有两个基因型,不能进行多重比较。分子标记Vsac05基因型间平均值的多重比较结果表明 (表6):含BC基因型个体的体长/体高平均值最小 (1.389),最大为AA等位基因型个体 (1.745);含有基因型AA个体的体长/体高及全长/体高平均值显著高于含AC、AD和BC 3种基因型的个体,而含BD基因型个体的体长/体高平均值显著高于含BC基因型的个体 (P<0.05),其他基因型个体体长/体高及全长/体高均无显著性差异 (P>0.05)。AC、AD、BC基因型只在YT家系中出现,AA和BD在两个家系中均有出现,等位基因C(375 bp)是圆头家系特有的基因。
3 讨论
3.1 血鹦鹉体型与分级挑选
在鱼类生产上,通常利用杂种优势选育具有生长快或抗逆性强等优良性状的品种[13-16]。观赏鱼在利用杂种优势进行选择时则更侧重于选择体型和体色。血鹦鹉的体型既不像父本也不像母本,作为杂交后代其性状不稳定,即使是全同胞家系,个体间的体型和体色也差异较大。血鹦鹉正是通过这种“杂种优势”选育出金刚鹦鹉、元宝鹦鹉、罗汉鹦鹉、麒麟鹦鹉和红财神等多个品种。尽管血鹦鹉雌鱼性成熟后可以正常排卵,雄鱼却未观察到完全能育型的精巢及正常排精的现象,所以血鹦鹉的优良性状较难稳定遗传[17]。因此,本研究中从血鹦鹉形态学性状和遗传多样性水平揭示不同体型血鹦鹉家系的种质差异,筛选与体长/体高相关的分子标记,以期为血鹦鹉亲本配组提供理论基础,提高血鹦鹉品种的优级率。
表6 标记Vsac05位点不同基因型性状平均值和多重比较
Tab.6 Means and multiple comparisons of traits with different genotypes in locus Vsac05

注:同列中标有不同小写字母表示组间有显著性差异 (P<0.05),标有相同小写字母表示组间无显著性差异 (P>0.05)
Note:The means with different letters within the same column are significant differences at the 0.05 probability level,and the means with the same letters within the same column are not significant differences
基因型gene type个体数/ind.number体长/体高body length/body height全长/体高total length/body height AA(337/337) 21 1.745±0.311a 2.350±0.341a AC(337/375) 6 1.400±0.105bc 2.001±0.141b AD(337/391) 5 1.415±0.172bc 1.993±0.231b BC(359/375) 11 1.389±0.084c 1.972±0.125b BD(359/391) 17 1.648±0.280ab 2.222±0.294ab
何丽等[17]对血鹦鹉体型各性状间的比例关系进行研究,发现体高/体长值越接近黄金比例(0.618)的血鹦鹉,观赏价值越高。然而,随着人们的审美标准及观赏需求的变化,目前,体长/体高越接近1.0的血鹦鹉品级越高,价格也越贵。按照血鹦鹉的国标分级挑选标准,血鹦鹉按照体长/体高可以分为AAAA级、AAA级、AA级、A级和B级鱼[18]。本研究中YT家系的体长/体高平均值为1.359,比值为1.1~1.2的个体占样本数的3.3%,1.2~1.3的个体占样本数的 20.0%,1.3~1.4的个体占样本数的53.3%,1.4~1.5的个体占样本数的23.3%,以上个体按照分级挑选标准分别属于AAA级、AA级、A级和B级鱼,均可作为商品鱼继续培养。JT家系的体长/体高平均值为1.795,比值大于1.5的个体为28尾,占样本数的93.3%,基本上均属于淘汰鱼。性状测量结果表明,YT家系的性状显著优于JT家系。本研究中利用SPSS 21.0软件对两个家系进行判别分析,得到的判别公式能较好地区分两个家系,综合判别率高达95%,可以为血鹦鹉分级挑选提供参考。
3.2 血鹦鹉家系的遗传多样性水平
基因决定性状,基因差异是个体性状差异的主要根源。微卫星标记是目前进行动物分子辅助育种研究最常用的分子标记之一[19-21]。遗传多样性水平高低是评价种质资源的重要指标,等位基因数、有效等位基因数、期望杂合度、观测杂合度和多态信息含量等均为反映群体遗传多样性水平高低的参数,这些参数值越大,遗传多样性水平越高,遗传变异越丰富。本研究中采用6个微卫星标记,除标记Vsac04和Vsac06以外,其他标记均已被证实处于中高等态多态水平且扩增良好[22],能够用于血鹦鹉遗传多样性分析及种质资源评估。固定系数(Fis)可反映观测杂合度和期望杂合度间的平衡关系,Fis越接近于零,基因型的分布就越接近平衡状态,Fis>0表明杂合子缺失,Fis<0表明杂合子过剩。本研究结果显示,两个家系均表现为杂合子过剩 (Fis<0)。综上所述,YT家系等位基因数、有效等位基因数、期望杂合度、观测杂合度、多态信息含量和香农信息指数的平均值均高于JT家系,表明圆头家系的遗传多样性水平较高,变异较丰富。
3.3 微卫星标记与性状的连锁分析
分子标记与性状的连锁分析就是对个体的位点基因型与表型性状进行显著性检验,差异显著则说明标记与性状的基因存在连锁遗传,在育种过程中可通过筛选性状相关的分子标记来缩短育种时间和提高选种准确性[6]。刘肖莲等[6]通过标记连锁分析发现,标记Vsac05和Vsac06与体长极显著相关,标记Vsac02与体高/体长极显著相关。在养殖生产中,体长/体高是影响血鹦鹉分级标准和价格的最重要因素,因此,本研究中采用SPSS 21.0软件的GLM模型,对微卫星标记和血鹦鹉体长/体高、全长/体高两个特征值进行关联分析,结果表明,标记Vsac04和Vsac05与体长/体高、全长/体高显著相关。关联分析表明,一个性状可以受多个基因控制,或者一个基因位点可以影响多个性状的表达。这一点与刘肖莲等[6]试验结果相同,即在性状关联分析中存在 “多因一效”和 “一因多效”现象。但由于所选试验群体不同,不同群体的遗传背景不同,因此,群体遗传多样性和关联分析的结果也不完全相同。对分子标记Vsac05进行多重比较表明,其基因型AC、AD、BC只在YT家系中出现,在育种实践中可以作为优先选择的基因型,等位基因C(375 bp)是圆头家系特有的基因。含等位基因C(375 bp)个体的体长/体高平均值低于其他基因型个体。在血鹦鹉进行育种时,体长/体高值越小 (比值越接近于1),圆头性状越明显,血鹦鹉的等级和价格越高。因此,分子标记Vsac05可用于血鹦鹉育种,在血鹦鹉亲本进行组配时,优先选择含有标记Vsac05等位基因C(375 bp)的父母本,使圆头优势性状得到保存和富集,从而提高血鹦鹉鱼的优级率。
综上所述,血鹦鹉体型受多个基因的遗传控制,遗传机制比较复杂。目前,关于血鹦鹉的分子研究尚少,所用的遗传标记更是寥寥无几。采用高通量测序技术和生物信息学的方法可以获得更多与生长调控相关的基因和分子标记。因此,还有许多工作亟需进一步开展,比如对血鹦鹉分子标记的开发、生长调控和褪色机制研究等。
参考文献:
[1] 孙志景,姜巨峰,傅志茹,等.红头丽体鱼×红魔丽体鱼杂交子一代胚胎发育及仔鱼形态学观察[J].南方水产科学,2014,10(3):38-46.
[2] 吴会民,姜巨峰,刘肖莲,等.陶粒在血鹦鹉温室大棚养殖水处理中的应用[J].天津农业科学,2016,22(4):36-39.
[3] 施振宁.杂交观赏鱼血鹦鹉的繁育与养殖技术[J].科学养鱼,2016(10):79-80.
[4] 崔培,范泽,李建,等.饲料中添加白藜芦醇对血鹦鹉肝脏生化指标的影响[J].大连海洋大学学报,2015,30(2):203-206.
[5] 夏苏东,傅志茹,谢刚,等.芦荟粉对血鹦鹉幼鱼体色与生长的影响[J].水产科学,2016,35(1):7-13.
[6] 刘肖莲,姜巨峰,吴会民,等.血鹦鹉遗传多样性及经济性状的关联分析[J].西北农林科技大学学报:自然科学版,2016,44(12):51-55,63.
[7] Xu P,Zhang X F,Wang X M,et al.Genome sequence and genetic diversity of the common carp,Cyprinus carpio[J].Nature Genetics,2014,46(11):1212-1219.
[8] Jin S B,Zhang X F,Jia Z Y,et al.Genetic linkage mapping and genetic analysis of QTL related to eye cross and eye diameter in common carp(Cyprinus carpio L.)using microsatellites and SNPs[J].Aquaculture,2012,358-359:176-182.
[9] Kuang Y Y,Zheng X H,Lv W H,et al.Mapping quantitative trait loci for flesh fat content in common carp(Cyprinus carpio)[J].Aquaculture,2015,435:100-105.
[10] 刘永新,刘奕,周勤,等.利用微卫星标记指导红鳍东方鲀亲本选配[J].大连海洋大学学报,2015,30(2):113-119.
[11] 汪金海,韩松,郑小东.金乌贼(Sepia esculenta)繁殖模式的分子学鉴定[J].海洋与湖沼,2017,48(1):184-189.
[12] Kuang Y Y,Zheng X H,Li C Y,et al.The genetic map of goldfish(Carassius auratus)provided insights to the divergent genome evolutions in the Cyprinidae family[J].Scientific Reports,2016,6:34849.
[13] 夏德全,曹萤,杨弘,等.罗非鱼杂交F1代与亲本的遗传关系及其杂种优势的利用[J].中国水产科学,1999,6(4):29-32.
[14] 余勇,李琪,于红,等.长牡蛎中国群体和日本群体杂交子代的杂种优势分析[J].中国海洋大学学报:自然科学版,2016,46(2):35-41.
[15] 朱华平,卢迈新,黄樟翰,等.鱼类遗传改良研究综述[J].中国水产科学,2010,17(1):168-181.
[16] 孙谦,段强,桑士田,等.菲律宾蛤仔选育家系间杂交的Kung育种值及配合力分析[J].大连海洋大学学报,2017,32(4):381-386..
[17] 何丽,陈再忠.血鹦鹉的体型及生殖潜力评价[J].水产科技情报,2008,35(5):245-247.
[18] 中华人民共和国国家质量监督检验检疫总局.GB/T 30946-2014观赏鱼分级规则血鹦鹉鱼[S].北京:中国标准出版社,2015.
[19] 鲁翠云,曹顶臣,孙效文,等.微卫星分子标记辅助镜鲤家系构建[J].中国水产科学,2008,15(6):893-901.
[20] 戴欢,刘洋,王文文,等.半滑舌鳎抗哈维氏弧菌病相关微卫星标记筛选及 QTL 定位[J].中国水产科学,2017,24(1):22-30.
[21] Sun X W,Liang L Q.A genetic linkage map of common carp(Cyprinus carpio L.)and mapping of a locus associated with cold tolerance[J].Aquaculture,2004,238(1-4):165-172.
[22] Botstein D,White R L,Skolnick M,et al.Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J].American Journal of Human Genetics,1980,32(3):314-331.