摘要:为探明黄河鲇Silurus lanzhouensis幼鱼对饲料钙、磷的适宜需要量,选取总能、钙、粗蛋白质、非植酸磷、赖氨酸5个因素作为试验因素,每个因素各设6个水平,钙为0.50%~1.34%,非植酸磷为0.50%~1.00%,采用均匀设计法U*6(64)设计6种配合饲料,饲喂体质量为(170±5)g的黄河鲇幼鱼,分别记为A、B、C、D、E、F组,饲养时间为60 d。试验结束后,以增重率、特定生长率、肌肉和肝脏RNA/DNA值、粗蛋白质离体消化率为试验指标,采用偏最小二乘法建立回归方程,通过降维分析和优化求解,分析研究黄河鲇幼鱼对饲料中钙和非植酸磷的适宜需要量。结果表明:C组(钙0.56%、非植酸磷0.60%)试验鱼的增重率、特定生长率和肌肉RNA/DNA最高,分别为84.83%、1.02%和2.98,肠、肝、胃中粗蛋白质离体消化率也达到最大值,分别为67.53%、66.76%、51.52%;随着饲料中钙、非植酸磷含量的增加,鱼体增重率、特定生长率、肌肉RNA/DNA值和粗蛋白质离体消化率均呈现先上升后下降的趋势。研究表明,饲料中钙和非植酸磷含量对黄河鲇幼鱼生长、蛋白质合成能力、饲料粗蛋白质离体消化率影响明显,黄河鲇幼鱼对饲料中钙的适宜需要量为0.54%,对非植酸磷的适宜需要量为0.50%~0.53%。
关键词:黄河鲇;营养需求;钙;磷;均匀设计
钙和磷是鱼体内含量最多的无机元素,是维持鱼类骨骼发育及保证其正常生长的重要营养元素,且在维持酸碱平衡中起着重要作用[1],因此,饲料中钙磷含量对鱼类健康、生长和饲料利用率且有较大的影响[1-2]。鱼类可从养殖水体中吸收钙满足其生理需求[2-3],但由于自然水体中磷的浓度较低,且鱼类对水中磷的吸收能力较差,无法从水环境中吸收足够的磷,因此,饲料中磷的含量成为影响鱼类生长的重要因素之一[4]。
黄河鲇Silurus lanzhouensis俗称绵鱼,学名兰州鲇,隶属于鲇形目、鲇科、鲇属,全身光滑无鳞,皮肤富黏液,有较高的经济价值和营养价值[5]。近些年,由于野生种群的数量不断减少,黄河鲇已被《中国物种红色名录》列为濒危物种[6]。2004年,吴旭东等[5]在黄河宁夏段采集到一批黄河鲇标本,为宁夏一新纪录种。随后,邱小琮等[7]测定了黄河鲇肌肉的生化成分,并对其营养成分进行了评价,研究了黄河鲇对4种饲料原料的离体消化率和酶解能力[8],赛清云等[9]通过研究黄河鲇对饲料总能、粗蛋白质的需要,为研发配合饲料提供了一定的理论依据。目前,关于黄河鲇对饲料中钙磷的需求研究尚未见报道。本研究中,采用均匀设计的试验方法,配制了6种不同配方的营养饲料饲喂黄河鲇,测定了黄河鲇的增重率、特定生长率、鱼体肌肉与肝脏的RNA/DNA值,以及离体条件下黄河鲇对6组不同配方饲料的粗蛋白质和干物质消化率,通过建立回归方程和优化求解,分析了黄河鲇对非植酸磷、钙的适宜需求量,以期为黄河鲇饲料的合理配制提供科学依据,促进黄河鲇养殖业的健康发展。
1.1 材料
试验鱼为宁夏水产研究所提供的同一批黄河鲇1龄幼鱼,从中挑选出规格一致(170 g±5 g)、体格健壮的鱼用于试验。
1.2 方法
1.2.1 试验设计 试验选择赖氨酸、钙、粗蛋白质、总能、非植酸磷5个因素,每个因素设6个水平,饲料配方采用均匀设计方法![]() 设计(表1)。
设计(表1)。
表1 试验因素及水平在均匀设计表中的安排
Tab.1 Experimental factors and levels in uniform design table
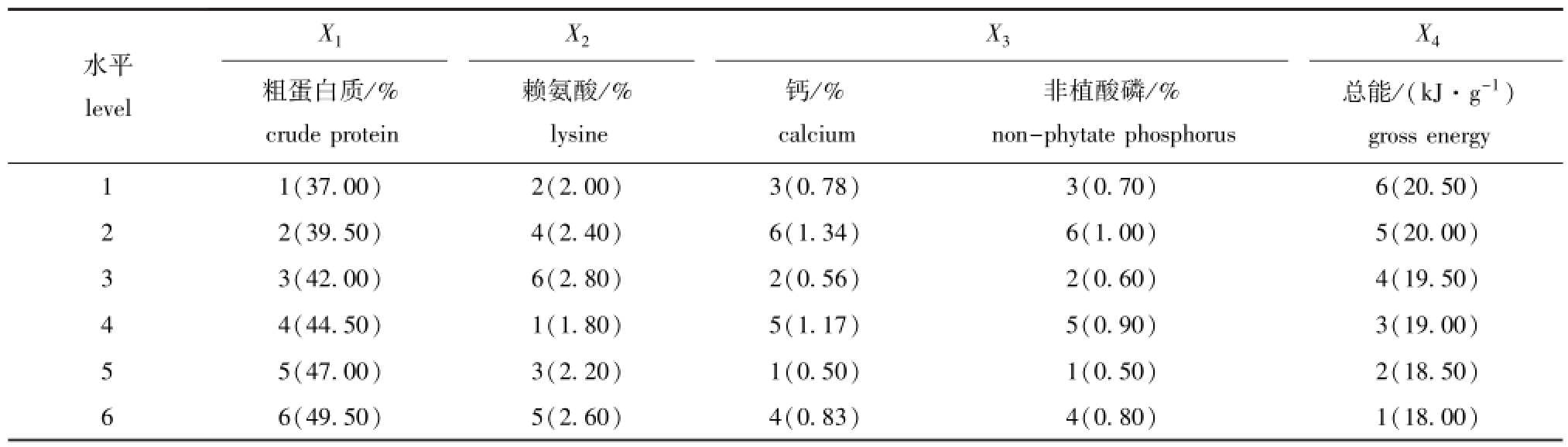
X1X2X3 X4水平level总能/(kJ·g-1) gross energy 1 1(37.00)2(2.00)3(0.78)3(0.70)6(20.50)粗蛋白质/% crude protein赖氨酸/% lysine钙/% calcium非植酸磷/% non-phytate phosphorus 2 2(39.50)4(2.40)6(1.34)6(1.00)5(20.00) 3 3(42.00)6(2.80)2(0.56)2(0.60)4(19.50) 4 4(44.50)1(1.80)5(1.17)5(0.90)3(19.00) 5 5(47.00)3(2.20)1(0.50)1(0.50)2(18.50) 6 6(49.50)5(2.60)4(0.83)4(0.80)1(18.00)
1.2.2 试验饲料的成分及测定
选取进口的酪蛋白、玉米蛋白粉、鱼粉、大豆粕作为主要蛋白源,能量水平通过豆油调节,用磷酸二氢钙调节磷和钙的含量,试验原料经超微粉碎处理,通过60目筛,依照配方进行称重、混匀,最终制成6种团状饲料,分别记为A、B、C、D、E、F,营养成分详见表2。本试验中,采用以二胺四乙酸络合快速滴定法(GB/T6436—2002)测定饲料中的钙含量,采用钼黄显色光度法(GB/ T6437—2002)测定磷含量。
6组饲料中钙含量的设计依照一定的钙磷比添加,在均匀设计表中钙含量的变化与非植酸磷的变化一致,因此,在数据分析时,可用钙代替非植酸磷,安排在因素X3中进行分析。
表2 试验饲料营养成分水平
Tab.2 The nutritional level of the experimental diets
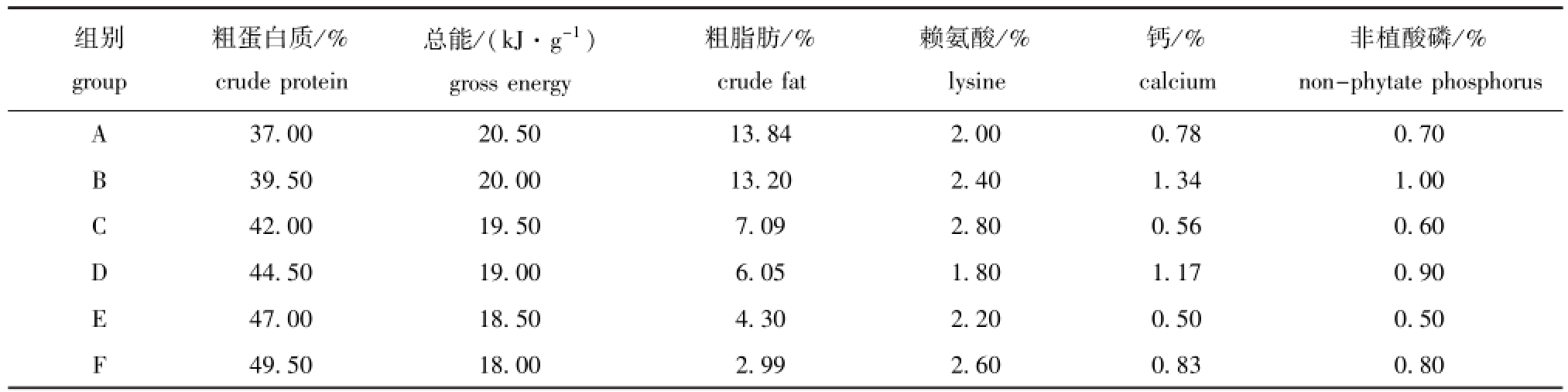
组别group粗蛋白质/% crude protein总能/(kJ·g-1) gross energy粗脂肪/% crude fat赖氨酸/% lysine钙/% calcium非植酸磷/% non-phytate phosphorus A 37.0020.5013.842.000.780.70 B 39.5020.0013.202.401.341.00 C 42.0019.507.092.800.560.60 D 44.5019.006.051.801.170.90 E 47.0018.504.302.200.500.50 F 49.5018.002.992.600.830.80
1.2.3 饲养与管理 将试验鱼饲养于网箱(2.0 m×1.5 m×1.2 m)中,试验前先用粉碎的新鲜羊肝投喂,待试验鱼正常摄食后,将人工配合饲料添加到羊肝中饱食投喂,并逐量增加配合饲料的比例,先按5∶1的比例混合,再增加到3∶1,驯化20 d后,完全用配合饲料代替羊肝。待试验鱼能正常摄食配合饲料后,将其分为6组,分别记为A、B、C、D、E、F组,每组设3个平行,共用18个网箱,每个网箱放20尾鱼,准确称重并测量其体长,作为试验鱼初始记录。试验期间,水温为24~28℃,pH为8.1,溶氧>4.5 mg/L,氨氮<0.5 mg/L,每天于6:00、18:00饱食投喂。投饲量可视摄食和天气情况做适当调整,试验周期为60 d。饲养期间及时清除粪便和残饵,以保持水质清新,并定期用高锰酸钾进行全池消毒。
1.2.4 样品的制备及指标的测定
(1)生长指标。在生长试验开始和结束时,均在禁投喂24 h后,测定各组黄河鲇的初始体质量、终末体质量,并计算增重率和特定生长率。计算公式为[11]
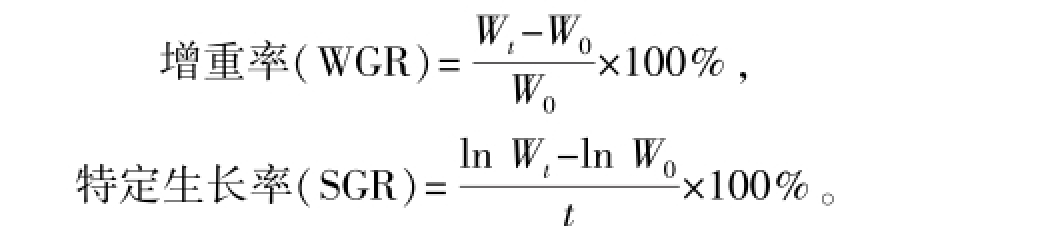
其中:Wt、W0分别为终末和初始鱼体总质量(g);t为试验时间(d)。
(2)肌肉和肝中的RNA/DNA值。在饲养试验结束后,将试验鱼饥饿24 h,从每组中取5尾鱼进行解剖,分别取适量肝组织及鱼背鳍基部白肌于液氮(-196℃)中研磨。从每个样品中快速准确称取30 mg和80 mg研磨组织各1份,保存于冰箱中待测。
参照Sambrook等[12]中的方法,用酚-氯仿法提取DNA。
采用AXyPrep总RNA小量制备试剂盒(美国AXygen公司)提取RNA,然后利用微量紫外分光光度计(Nano Drop ND-1000型)分别测定RNA和DNA含量,并计算RNA/DNA值。
每个处理组需要进行3次取样,将每一样品进行3次重复测定,然后取其平均值。
(3)粗蛋白质的离体消化率。
a.粗酶液的提取。参照文献[13]中的方法制备粗酶提取液。试验结束后,从所有试验网箱中随机捞出18尾饥饿24 h的试验鱼(170 g左右),随机分为6组,将各组黄河鲇的肝脏、胃和肠道分别取出,除去肠道内容物、胃、系膜和肠道脂肪,分别用滤纸吸干后称取肝、胃、肠的质量,然后按照1 g组织加入10 mL溶液的比例,分别加入磷酸缓冲液(0.2 mol/L,pH 7.4),用冰冻玻璃匀浆器迅速匀浆后,以4000 r/min离心30 min(在低温条件下),缓慢吸取上清液,将其作为粗酶提取液,于冰箱(-18℃)中保存备用,并在24 h内分析完成。
b.离体消化、酶解。准确称取6种配合饲料各0.4 g,放入100 mL的带塞三角瓶中,并加入40 mL 0.2 mol/L的磷酸缓冲液(pH 7.4),各组消化道各部位的粗酶液为10 mL,并将每种饲料设置3个平行;对照组则选用消化道各部位的煮沸粗酶液。然后加入150 IU/mL的硫酸链霉素和青霉素,置于(28±1)℃恒温生化培养箱中,并加以摇床振荡(50 r/min),对其进行离体消化、酶解。
c.离体消化率的测定。8 h后终止离体消化反应,定量滤纸过滤,取滤渣(含滤纸)在105℃下烘至恒重并称重。以凯氏定氮法测定6种饲料消化后相应滤渣的粗蛋白质含量,计算公式为
粗蛋白质消化率=(饲料样品粗蛋白质含量-消化后滤渣粗蛋白质含量)/饲料样品粗蛋白质含量×100%。
1.3 数据处理
试验数据使用DPS 14.50软件进行方差分析,建立二次多项式回归方程,采用偏最小二乘法对回归方程进行优化求解,以寻求试验因素间的最优组合;对所得二次多项式回归方程进行降维分析,得到试验因素与各指标的一元回归方程,并对降维方程作图分析,寻找单因素与所测指标的直接关系。
2.1 生长指标
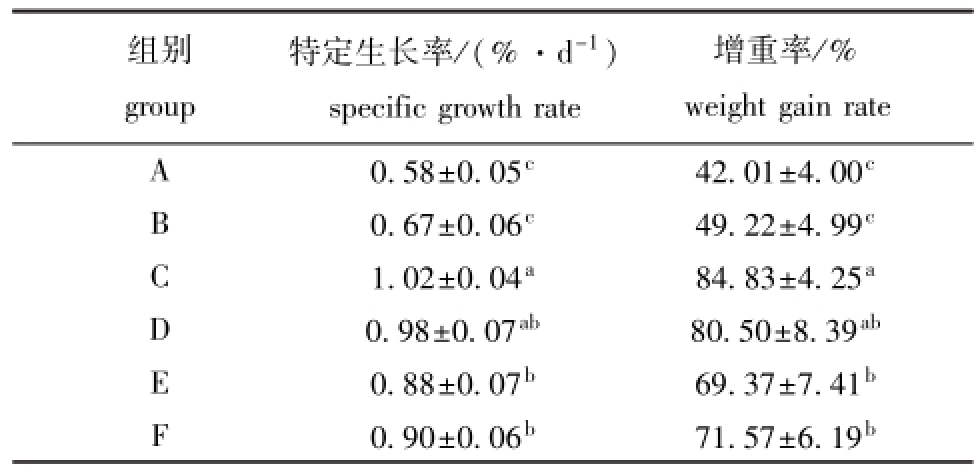
经过60 d的饲养试验,各试验组鱼的特定生长率和增重率结果见表3,摄食不同营养水平饲料的黄河鲇增重率和特定生长率有显著性差异(P<0.05)。C组黄河鲇幼鱼的特定生长率和增重率均最高,分别为1.02%/d、84.83%,且均显著高于A、B、E、F组(P<0.05),与D组无显著性差异(P>0.05)。
表3 摄食不同配方饲料时黄河鲇的生长指标
Tab.3 Growth of catfishSilurus lanzhouensisfed different diets
注:同列中标有不同小写字母者表示组间有显著性差异(P<0.05),标有相同小写字母者表示组间无显著性差异(P>0.05),下同
Note:The means with different letters within the same column are significant differences at the 0.05 probability level,and the means with the same letters within the same column are not significant differences at the 0.05 probability level,et sequentia
组别group增重率/% weight gain rate A 0.58±0.05c42.01±4.00特定生长率/(%·d-1) specific growth rate c 0.67±0.06c49.22±4.99cC 1.02±0.04a84.83±4.25aD 0.98±0.07ab80.50±8.39abE 0.88±0.07b69.37±7.41bF 0.90±0.06b71.57±6.19 B b
2.2 各组鱼肌肉和肝中的RNA/DNA值
从表4可见:摄食不同营养水平饲料的黄河鲇鱼体RNA/DNA值有显著性差异(P<0.05);C组鱼肌肉的RNA/DNA值最大,且显著高于A、B、F组(P<0.05),D组鱼肝脏的RNA/DNA值最大,且显著高于A、B组(P<0.05)。
2.3 黄河鲇对各组试验饲料蛋白质的离体消化率
从表5可见:蛋白质水平最高的F组,其粗蛋白质消化率并不高,而C组鱼肠、肝的粗蛋白质消化率均最大,均显著高于其他饲料组(P<0.05),C组鱼胃的粗蛋白质消化率也最大,除与D组无显著性差异(P>0.05)外,均显著高于其他饲料组(P<0.05);蛋白质水平最低的A组,其肠、肝、胃的粗蛋白质消化率均最低。
表4 摄食不同配方饲料的黄河鲇肌肉和肝中的RNA/DNA值
Tab.4 RNA/DNA values in muscle and liver of catfishSilurus lanzhouensisfed different diets
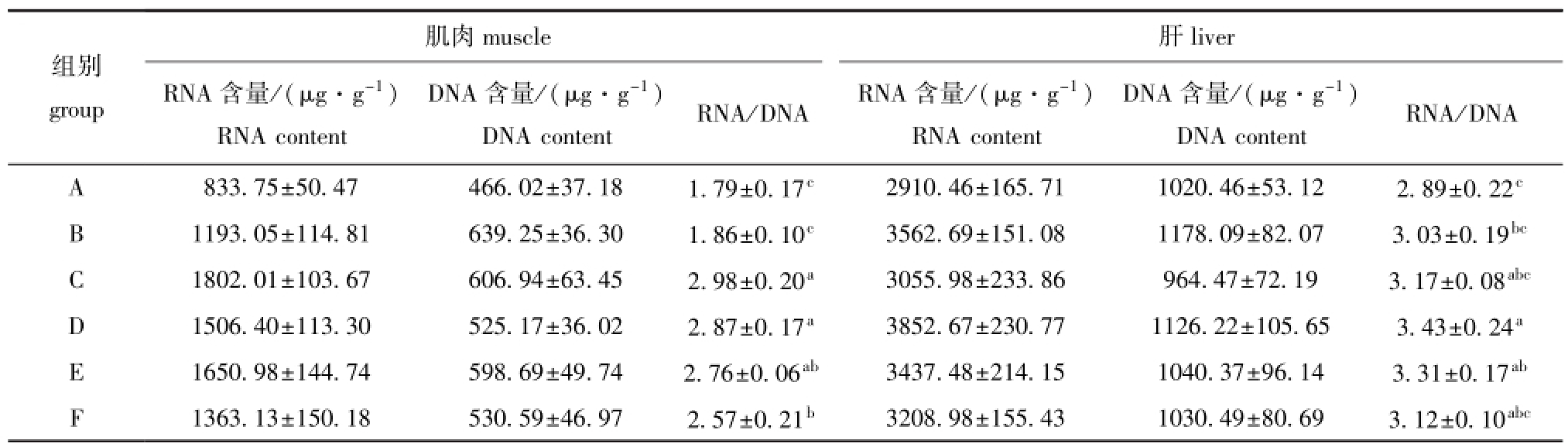
muscle肌肉组别group肝liver RNA含量/(μg·g-1) RNA content DNA含量/(μg·g-1) DNA contentRNA/DNARNA含量/(μg·g-1) RNA content DNA含量/(μg·g-1) DNA content RNA/DNA A 833.75±50.47466.02±37.181.79±0.17c2910.46±165.711020.46±53.122.89±0.22cB1193.05±114.81639.25±36.301.86±0.10c3562.69±151.081178.09±82.073.03±0.19bcC1802.01±103.67606.94±63.452.98±0.20a3055.98±233.86964.47±72.193.17±0.08abcD1506.40±113.30525.17±36.022.87±0.17a3852.67±230.771126.22±105.653.43±0.24aE1650.98±144.74598.69±49.742.76±0.06ab3437.48±214.151040.37±96.143.31±0.17abF1363.13±150.18530.59±46.972.57±0.21b3208.98±155.431030.49±80.693.12±0.10abc
表5 黄河鲇对饲料蛋白质的离体消化率
Tab.5 The in vitro digestibility of crude protein in test diets by catfishSilurus lanzhouensis%
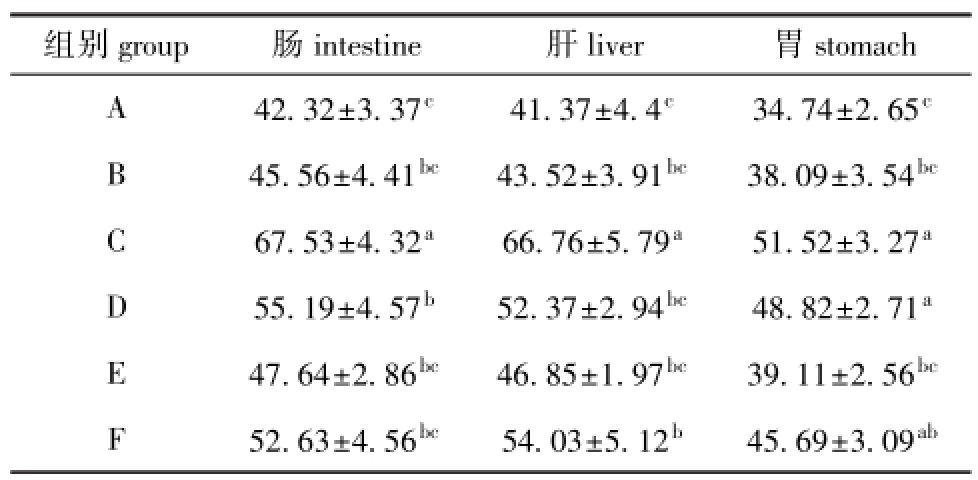
组别group肠intestine肝liver胃stomach A42.32±3.37c41.37±4.4c34.74±2.65cB45.56±4.41bc43.52±3.91bc38.09±3.54bcC67.53±4.32a66.76±5.79a51.52±3.27aD55.19±4.57b52.37±2.94bc48.82±2.71aE47.64±2.86bc46.85±1.97bc39.11±2.56bcF52.63±4.56bc54.03±5.12b45.69±3.09ab
2.4 回归方程的建立及其降维分析与优化求解
2.4.1 生长指标与饲料钙、非植酸磷的关系 采用偏最小二乘回归法,以粗蛋白质含量(X1)、赖氨酸含量(X2)、钙含量(X3)、总能含量(X4)为自变量,以相对增重率(Y11)、特定生长率(Y12)为因变量,建立回归方程如下:

回归方程中,Y11、Y12的决定系数R2分别为0.990 7、0.995 7,经F检验P<0.05,表明回归方程的拟合程度较好。
采用偏最小二乘回归法,以粗蛋白质含量(X1)、赖氨酸含量(X2)、非植酸磷含量(X3)、总能(X4)为自变量,以相对增重率(Y13)、特定生长率(Y14)为因变量,建立回归方程如下:

回归方程中,Y13、Y14的决定系数R2分别为0.991 6、0.990 7,经F检验P<0.05,表明回归方程的拟合程度较好。
将其他因子固定为1水平,由Y11、Y12得到相对增重率、特定生长率与钙的一元回归方程为
将其他因子固定为1水平,由Y13、Y14得到相对增长率、特定生长率与非植酸磷的一元回归方程为
对降维方程作图,由图1~图2可知,随着饲料中钙、非植酸磷含量的增加,相对增重率与特定生长率均呈先上升后下降趋势。
在本试验条件下,为了寻求黄河鲇增重率和特定生长率的最大值,利用DPS数据处理系统对回归方程进行了优化求解。结果显示:当钙为0.54%时,最优目标函数Y11(增重率)为79.30%,Y12(特定生长率)为0.96%/d;当非植酸磷为0.51%时,最优目标函数Y13(增重率)为85.31%,Y14(特定生长率)为1.02%/d。
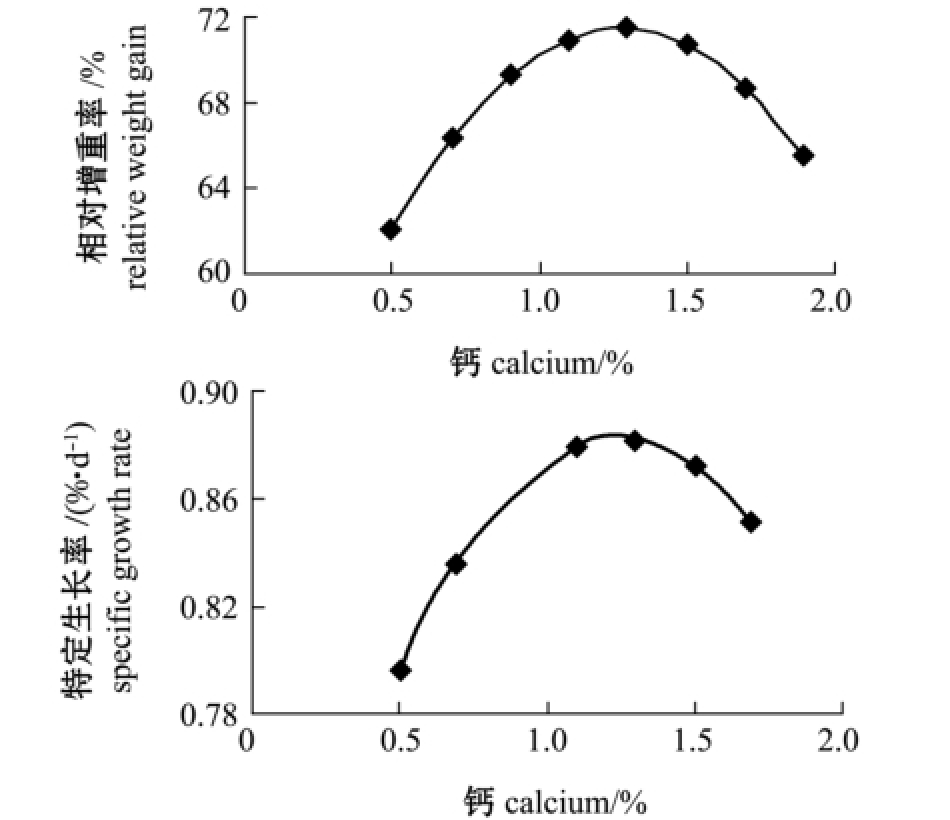
图1 钙与相对增重率、特定生长率的变化趋势图
Fig.1 Relationship between relative weight gain rate and specific growth rate and dietary calcium level
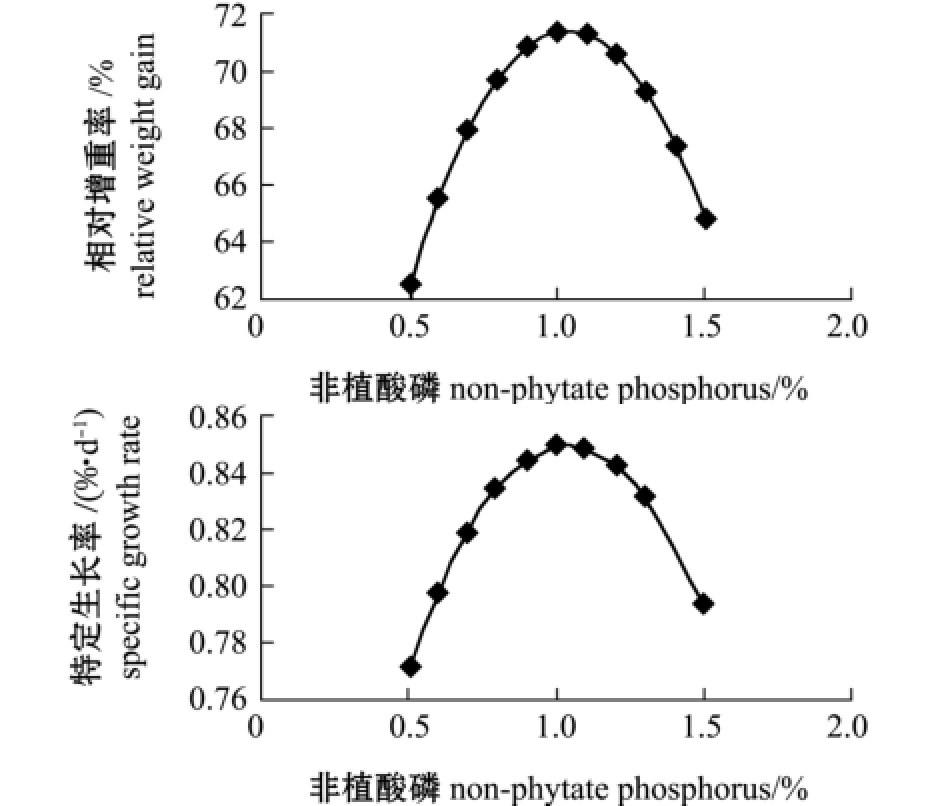
图2 非植酸磷与相对增重率、特定生长率的变化趋势图
Fig.2 Relationship between dietary non-phytate phosphorus level and relative weight gain rate and specific growth rate
2.4.2 黄河鲇肌肉和肝中的RNA/DNA值与饲料中钙、非植酸磷的关系 采用偏最小二乘回归法,以粗蛋白质含量(X1)、赖氨酸含量(X2)、钙含量(X3)、总能含量(X4)为自变量,以黄河鲇肌肉的RNA/DNA(Y21)、肝的RNA/DNA(Y22)为因变量,建立回归方程如下:


回归方程中,Y21、Y22的决定系数R2分别为0.987 1、0.785 8,经F检验P<0.05,表明回归方程的拟合程度较好。
采用偏最小二乘回归法,以粗蛋白质含量(X1)、赖氨酸含量(X2)、非植酸磷含量(X3)、总能(X4)为自变量,以黄河鲇肌肉的RNA/DNA (Y23)、肝的RNA/DNA(Y24)为因变量,建立回归方程如下:

回归方程中,Y23、Y24的决定系数R2分别为0.975 9、0.801 8,经F检验P<0.05,表明回归方程的拟合程度较好。
将其他因子固定为1水平,由Y21、Y22得到黄河鲇肌肉、肝的RNA/DNA值与钙的一元回归方程为
将其他因子固定为1水平,由Y23、Y24得到黄河鲇肌肉、肝的RNA/DNA值与非植酸磷的一元回归方程为
对降维方程作图,由图3~图4可见:黄河鲇肌肉的RNA/DNA值随着饲料中非植酸磷含量的增加,呈先上升后下降的趋势,但肝脏中的RNA/ DNA值并无此趋势,曲线最高点靠近纵坐标,在试验中设计的非植酸磷水平范围内呈不断下降趋势;而随着钙水平的不断升高,肌肉和肝脏中RNA/DNA值逐渐升高,当到达最大值时,如果继续提高钙的含量,RNA/DNA又开始下降。
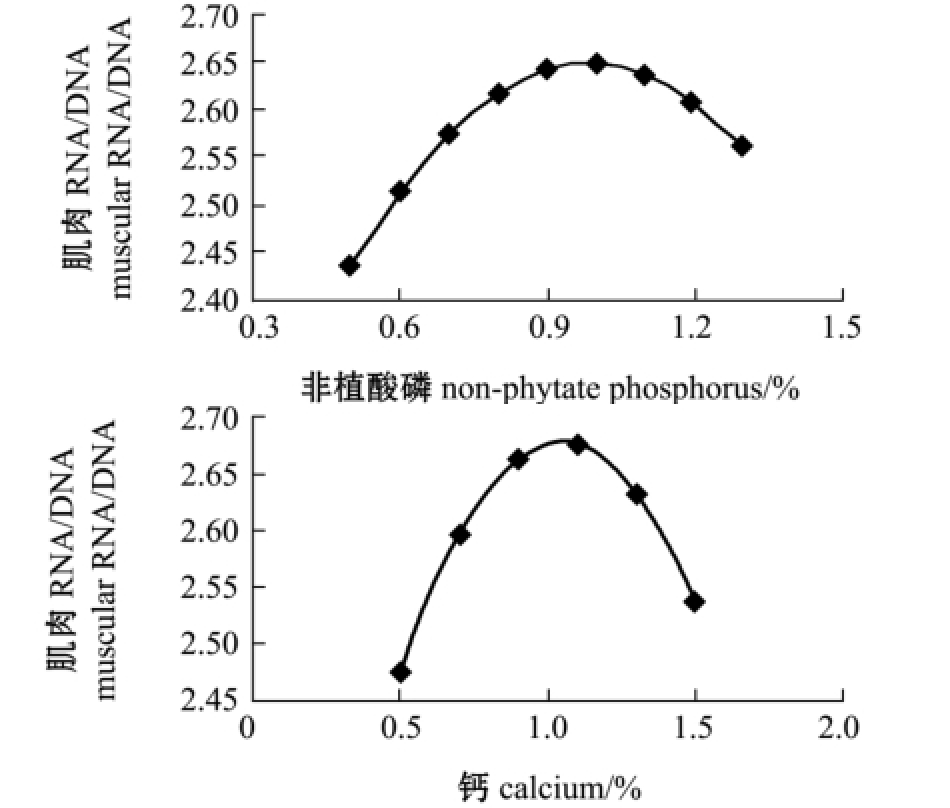
图3 钙、非植酸磷与肌肉中RNA/DNA值的变化趋势图
Fig.3 Relationship between dietary calcium level and non-phytate phosphorus level and RNA/DNA in muscle
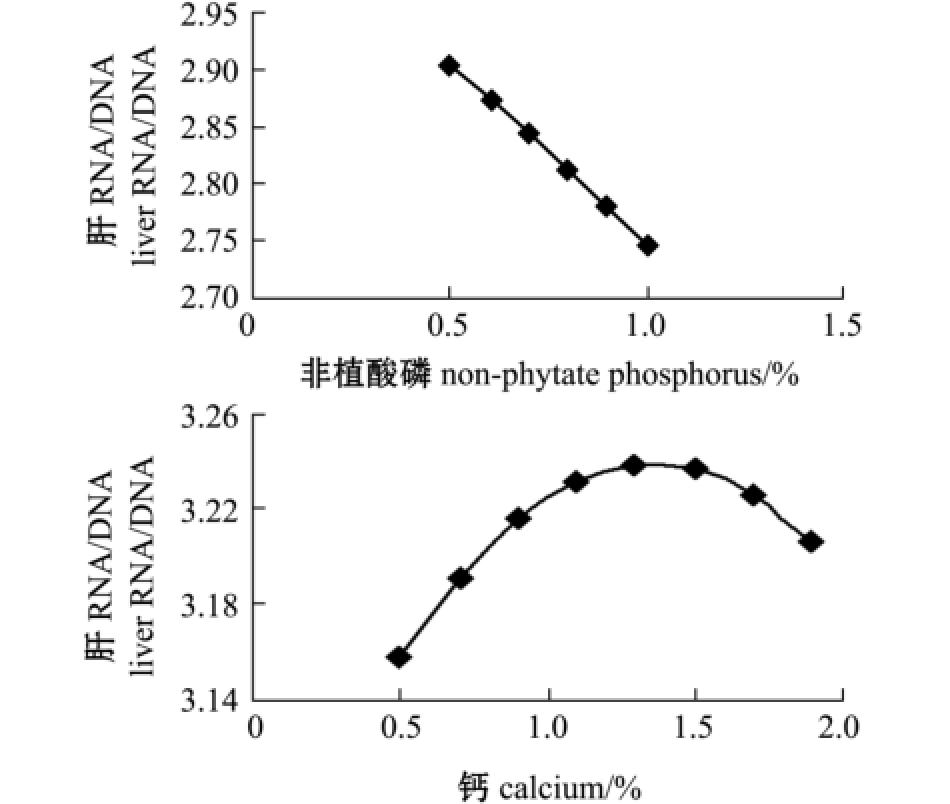
图4 钙、非植酸磷与肝脏中RNA/DNA值的变化趋势图
Fig.4 Relationship between dietary calcium level and non-phytate phosphorus level and RNA/DNA in the liver
在本试验条件下,为寻求黄河鲇肌肉和肝中RNA/DNA值的最大值,通过DPS数据处理系统对回归方程进行优化求解,得到各个试验因素的最优组合为:钙为0.54%时,最优目标函数Y21(肌肉RNA/DNA)为2.96,最优目标函数Y22(肝脏RNA/DNA)为3.34;非植酸磷为0.53%时,最优目标函数Y23(肌肉RNA/DNA)为3.05,最优目标函数Y24(肝脏RNA/DNA)为3.37。
2.4.3 粗蛋白质消化率与饲料钙、非植酸磷的关系 采用偏最小二乘回归法,以粗蛋白质含量(X1)、赖氨酸含量(X2)、钙含量(X3)、总能含量(X4)为自变量,以黄河鲇肠、肝、胃中粗蛋白质消化率为因变量,建立回归方程如下:

回归方程中,Y31、Y32、Y33的决定系数R2分别为0.960 5、0.984 2、0.909 6,经F检验P<0.05,表明回归方程的拟合程度较好。
采用偏最小二乘回归法,以粗蛋白质含量(X1)、赖氨酸含量(X2)、非植酸磷含量(X3)、总能含量(X4)为自变量,以黄河鲇肠、肝、胃中粗蛋白质消化率为因变量,建立回归方程如下:

回归方程中,Y34、Y35、Y36的决定系数R2分别为0.962 4、0.979 3、0.991 2,经F检验P<0.05,表明回归方程的拟合程度较好。
将其他因子固定为1水平,由Y31、Y32、Y33,得到黄河鲇肠、肝、胃中粗蛋白质消化率与钙的一元回归方程为

将其他因子固定为1水平,由Y34、Y35、Y36,得到黄河鲇肠、肝、胃中粗蛋白质消化率与非植酸磷的一元回归方程为

对所得到的降维方程作图,由图5~图6可见:黄河鲇肠、肝、胃中粗蛋白质的离体消化率随着饲料中钙含量的增加缓慢上升,当钙含量继续增加,粗蛋白质消化率反而有下降趋势;随着饲料中非植酸磷含量的不断增加,黄河鲇肠、肝、胃中粗蛋白质的离体消化率也呈明显的先上升后下降的趋势。
在本试验条件下,为寻求黄河鲇对饲料粗蛋白质消化率的最大值,通过DPS数据处理系统对回归方程进行优化求解可知:当钙为0.54%时,最优目标函数Y31(肠中粗蛋白质消化率)为65.22%,Y32(肝中粗蛋白质消化率)为64.99%, Y33(胃中粗蛋白质消化率)为52.33%;当非植酸磷为0.50%时,最优目标函数Y34(肠中粗蛋白质消化率)为66.57%,Y35(肝中粗蛋白质消化率)为65.75%,Y36(胃中粗蛋白质消化率)为53.69%。

图5 肠、肝和胃中粗蛋白质消化率与钙的关系
Fig.5 Relationship between the digestibility of crude protein and dietary calcium level in intestine,liver and stomach
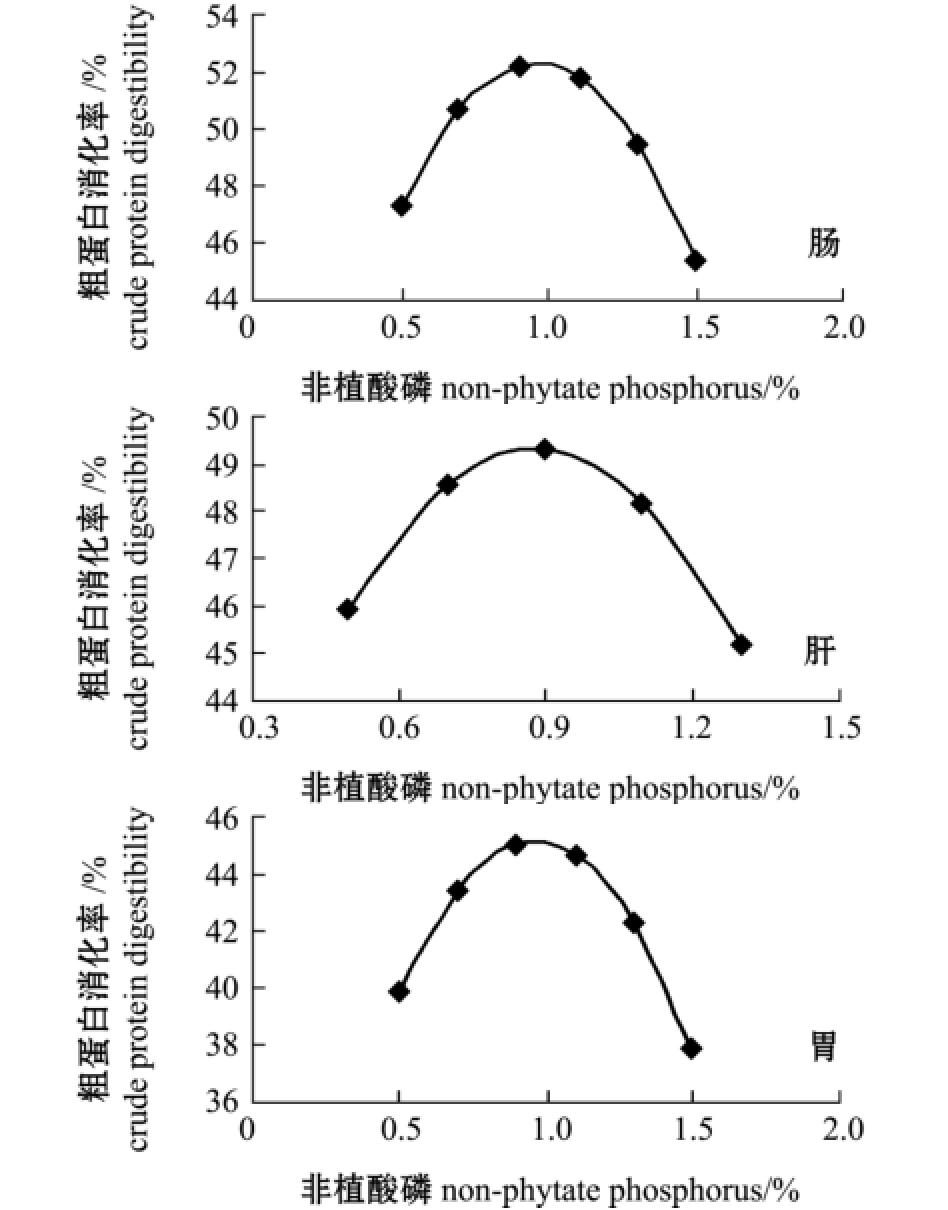
图6 肠、肝和胃中粗蛋白质消化率与非植酸磷的关系
Fig.6 Relationship between digestibility of crude protein and dietary non-phytate phosphorus level in intestine,liver and stomach
3.1 黄河鲇生长与钙、磷的关系
饲料中磷含量是影响鱼类生长的重要因素之一,目前,国内外有关鱼类对磷需要量的报道较多,但不同的鱼对磷需要量不尽相同。刘行彪等[14]以斑点叉尾鮰特定生长率为指标,通过折线模型分析发现,添加1435 U/kg植酸酶,等效于添加0.37%的有效磷,效果最佳;以椎骨磷为响应指标,通过折线模型分析发现,添加1226 U/kg植酸酶,等效于添加0.33%的有效磷,效果最佳。Lovell[15]研究表明,斑点叉尾鮰磷需要量为0.45%。陈建明等[16]研究表明,当饲料磷升高到0.88%时,翘嘴鲌鱼体增重和饲料效率最高,钙、镁、钾含量的变化对增重的影响均不显著,且饲料中若不添加这3种元素,鱼的增重率反而会相对较高,在生产性饲料中这3种元素含量一般较高,因此,在饲料中通常不需要添加。本研究结果表明,随着饲料中非植酸磷添加量的增加,增重率、特定生长率随之上升,但当其含量达到一定值时,继续增加非植酸磷含量,黄河鲇的生长开始缓慢下降。由此可见,磷是影响黄河鲇生长的重要因素之一,但饲料中磷含量不宜太高,过高反而会影响鱼的生长。这与对虹鳟[2,17]、草鱼[18]、镜鲤[19]的研究结果相似。
目前,关于鱼类饲料中是否应添加钙,意见并不一致。有研究表明,鱼类饲料中不必添加钙,鱼类从水体和基础饲料中获得的钙足以满足自身需要,饲料中添加钙不仅不会促进鱼类生长,反而会干扰鱼类对磷的吸收,影响鱼的正常生长和骨骼钙化[20];但也有研究表明,饲料中缺少钙会影响鱼类的正常生长,饲料中添加适量的钙能维持鱼类正常生长,促进磷的吸收[21]。本研究结果表明,随着饲料中钙含量的增加,增重率、特定生长率均呈先上升后下降的趋势,在钙为0.54%时,增重率和特定生长率均为最大值。
3.2 黄河鲇蛋白质合成(RNA/DNA)与钙、磷的关系
幼鱼消化系统发育不及成鱼完善,对饲料因子变化的反应较敏感,尤其是肉食性鱼类对饲料中蛋白要求较高[22]。蛋白质的合成对鱼类生长至关重要,RNA/DNA表示RNA的转录效率或蛋白质的合成或潜力核糖体的数量,也是衡量组织蛋白质合成能力的指标,RNA/DNA不仅与鱼体的生长有关,也与鱼类的营养状况有关[23]。肝脏是合成蛋白质的重要器官,白肌是蛋白质合成最终沉淀下来的最高组织[23],因此,肝脏、鱼肌肉的RNA/DNA值,不仅可以反映鱼体的蛋白质合成水平与合成状况[23],而且可以很好地反映鱼体的生长状况[9]。
本研究结果表明,在饲料中添加一定含量的钙、磷对黄河鲇的蛋白质合成(RNA/DNA)有明显的影响。随着饲料中钙和非植酸磷添加量的增加,RNA/DNA值也随之上升,当RNA/DNA值达到一定值时,继续增加钙和非植酸磷,RNA/DNA值开始缓慢下降,这与王桂芹等[23]对翘嘴鲌Culteralburnus、刘存歧等[24]对日本沼虾Macro-brachium nipponense、司亚东等[25]对鲤Cyprinus carpio的研究结果相似。饲料中添加适宜的钙、磷可促进黄河鲇蛋白质的合成,进而促进其生长。
3.3 离体消化率与钙、磷的关系
离体消化率是利用研究对象的消化道和消化器官酶提取液在试管内进行的消化试验,离体消化率是评价鱼类对饲料原料的可利用性的综合指标之一,其测定值可反映鱼对饲料的消化利用情况。离体消化率高则对营养物质的吸收能力强,有利于鱼体生长[26-27]。向枭等[28]认为,蛋白质品质以及饲料原料中的一些抗营养因子、日粮中必需氨基酸的平衡等因素,均会影响鱼对不同饲料干物质和粗蛋白质的消化率。
本研究结果表明,随着饲料中钙和非植酸磷含量的增加,黄河鲇对饲料粗蛋白质离体消化率呈先上升后下降的趋势,说明饲料中适宜钙、磷含量可促进黄河鲇对饲料粗蛋白质的离体消化率,但含量过高,则会抑制其离体消化率。
通过对方程优化求解和对试验指标的综合分析,黄河鲇对饲料非植酸磷最适需要量为0.50%~0.53%,此结果与斑点叉尾鮰对磷的需要量0.45%[15]相近,低于黑鲷(0.99%)、翘嘴鲌(0.88%)、异育银鲫(0.92%)[29]。黄河鲇对饲料钙的最适需要量为0.54%,此结果与宝石鲈对饲料钙的需要量(0.5%)[3]相近,低于赤眼鳟[20](0.6%~1.8%)。不同鱼类对饲料钙、磷的需要不同,这与鱼的种类、规格、年龄等因素有关,同时也受水质、水温等环境条件影响。
参考文献:
[1] 姚鹰飞,文华,蒋明,等.吉富罗非鱼饲料钙磷比研究[J].西南农业科技大学学报:自然科学版,2012,40(4):38-46,53.
[2] Ogino C,Takeda H.Mineral requirements in fish:Ⅲ.calcium and phosphorus requirements in carp[J].Bulletin of the Japanese Society of Scientific Fisheries,1976,42(7):793-799.
[3] 宋理平,冒树泉,王爱英,等.饲料钙磷水平对宝石鲈生长和体成分的影响[J].水产学报,2009,33(3):488-495.
[4] NRC(National Research Council).Nutrient Requirements of Fish and Shrimp[M].Washington,DC:National Academy Press,2011: 176.
[5] 吴旭东,张奇,赵红雪,等.宁夏鲇属鱼类一新纪录种——兰州鲇形态学特征描述[J].淡水渔业,2006,36(3):26-29.
[6] 汪松,解焱.中国物种红色名录:第1卷[M].北京:高等教育出版社,2004.
[7] 邱小琮,赵红雪,王远吉,等.兰州鲇肌肉营养成分分析及营养价值评价[J].水产科学,2008,27(8):407-410.
[8] 邱小琮,赵红雪,王远吉,等.兰州鲇对4种饲料原料的离体消化率和酶解能力[J].淡水渔业,2008,38(2):31-35.
[9] 赛清云,王远吉,吴旭东,等.黄河鲇幼鱼对饲料蛋白和能量需要的初步研究[J].淡水渔业,2012,42(4):53-58.
[10] 方开泰.均匀设计与均匀设计表[M].北京:科学出版社, 1994:53-69.
[11] 鲁宏申,石振广,魏德强,等.不同投饵率对达氏鳇幼鱼生长影响的初步研究[J].水生态学杂志,2010,3(1):147-149.
[12] Sambrook J F,Russell D W.Molecular Cloning:A Laboratory Manual[M].3rd.New York:Cold Spring Harbor Laboratory Press,2001:463-470.
[13] 叶元土,林仕梅,冯兴无,等.长吻鮠和南方大口鲶胃肠道消化能力的研究[J].动物学研究,1997,18(3):305-313.
[14] 刘行彪,黄可,付熊,等.植酸酶对斑点叉尾鮰生长性能及磷当量的研究[J].水生生物学报,2012,36(1):57-65.
[15] Lovell R T.Dietary phosphorus requirement of channel catfish (Ictalurus punctatus)[J].Transactions of the American Fisheries Society,1978,107(4):617-621.
[16] 陈建明,叶金云,潘茜,等.翘嘴鲌鱼种对磷的需求量[J].水生生物学报,2007,31(1):99-103.
[17] Coloso R M,King K,Fletcher J W,et al.Phosphorus utilization in rainbow trout(Oncorhynchus mykiss)fed practical diets and its consequences on effluent phosphorus levels[J].Aquaculture, 2003,220(1-4):801-820.
[18] 游文章,黄忠志,廖朝兴,等.草鱼对饲料中磷需要量的研究[J].水产学报,1987,11(4):285-292.
[19] Kim J D,Kim K S,Song J S,et al.Optimum level of dietary monocalcium phosphate based on growth and phosphorus excretion of mirror carp,Cyprinus carpio[J].Aquaculture,1998,161 (1-4):337-344.
[20] 郑惠芳,夏中生,黄凯,等.赤眼鳟鱼种配合饲料适宜钙磷水平及其比例的研究[J].淡水渔业,2007,37(5):45-49.
[21] Hossain M A,Furuichi M.Essentiality of dietary calcium supplement in redlip mullet Liza haematocheila[J].Aquaculture Nutrition,2000,6(1):33-38.
[22] 孟祥科,孙阳,屈菲,等.植酸酶对红鳍东方鲀幼鱼生长、消化酶及消化率的影响[J].大连海洋大学学报,2013,28(4): 323-328.
[23] 王桂芹,周洪琪,陈建明,等.饲料蛋白对翘嘴鲌蛋白质合成能力的影响[J].南京农业大学学报,2008,31(2):149-153.
[24] 刘存歧,沈会涛,吴玲玲.日本沼虾体内RNA/DNA比值与其生长关系的研究[J].河北大学学报:自然科学版,2006,26 (5):524-528.
[25] 司亚东,金有坤,周洪琪,等.鲤鱼白肌中RNA/DNA值与其生长的关系[J].上海水产大学学报,1992,1(3-4):159-167.
[26] 王友慧,叶元土,林仕梅,等.3种鱼对鱼粉酶解动力学及体外消化率的研究[J].西南农业大学学报,2002,24(3):259-262.
[27] 张玲,张卫东,张小玲,等.外源性蛋氨酸对鲤鱼离体消化率及酶解速度的影响[J].水生生物学报,2012,36(6):1200-1204.
[28] 向枭,叶元土,周兴华,等.铜鱼肠道、肝胰脏对四种蛋白质饲料的离体消化率测定[J].饲料工业,2003,24(1):50-51.
[29] 汤峥嵘,王道尊.异育银鲫和青鱼对饲料中钙、磷需要量的研究[J].上海水产大学学报,1998,7(S):140-147.
Requirements of juvenile catfish Silurus lanzhouensi for dietary calcium and non-phytate phosphorus
Abstract:The juvenile catfish Silurus lanzhouensis with body weight of(170±5)g were fed six formulated diets containing calcium from 0.50%to 1.34%,non-phytate phosphorus from 0.50%to 1.00%and six levels of gross energy,crude protein,and lysine with uniform design U*6(64)for 60 days to estimate the requirements of juvenile catfish for calcium and non-phytate phosphorus.The results showed that the maximum growth performance and ratio of RNA/DNA,and the maximum weight gain(84.83%),specific growth rate(1.02%),and ratio of RNA/ DNA(2.98)in muscle were observed in the fish fed the third diet containing 0.56%of calcuium,and 0.60%of non-phytate phosphorus.There were maximal in vitro apparent crude protein digestibility in intestine(67.53%), liver(66.76%)and stomach(51.52%),and weight gain,specific growth rate,and the ratio of RNA/DNA in muscle were increased with increase in the content of dietary calcium and non-phytate phosphorus at the early stage and then decreased in the late stage.The findings indicated that growth performance,protein synthesis capacity, and the crude protein in vitro digestibility were significantly influenced by dietary calcium and non-phytate phosphorus levels.It is recommended that the optimal dosage of calcium in diets be 0.54%,and that of non-phytate phosphorus ranged from 0.50%to 0.53%for juvenile catfish.
Key words:Silurus lanzhouensis;nutritional requirement;calcium;phosphorus;uniform design
DOI:10.16535/j.cnki.dlhyxb.2017.02.012
文章编号:2095-1388(2017)02-0189-09
中图分类号:S963.73;S965.1
文献标志码::A
收稿日期:2016-06-08
基金项目:宁夏科研院所专项(200906)