摘要:为研究自交对三疣梭子蟹Portunustrituberculatus幼体发育和形态性状的影响,通过人工定向交尾的方式,以建立的三疣梭子蟹连续1~9代家系内自交全同胞家系为材料,评价连续多代自交幼体发育和形态性状的影响。结果表明:在幼体发育方面,F1~F9代的排幼量无明显上升或下降趋势,但变态率研究中发现,溞状幼体和大眼幼体2个阶段中总体呈现衰退趋势,尤其自交家系程度高的F9代变态率最低,F9代Z1~M的4个阶段中变态率分别为74.87%、73.50%、59.17%、41.96%;在形态学方面,随着生长日龄与自交代数的增加,F3~F9代各项指标均低于F1、F2代家系,其中F9代各项指标皆为最小且与其他各代有显著性差异(P<0.05),但在150日龄时对梭子蟹整齐度的研究发现,F9代甲宽、甲长、体高变异系数分别为7.6365、7.5591、8.3282,在所有家系中为最小,整齐度最好;除F2代家系外,随着姊妹交系数的增加,相对于F1代,全甲宽出现-6.12%~-21.95%的衰退,甲宽衰退系数为-1.85%~-16.36%,甲长衰退系数为-10.08%~-27.66%,体高衰退系数为-9.39%~-22.70%,体质量衰退系数为-13.39%~-47.99%,排幼量衰退系数为-9.03%~-45.13%。研究表明,自交对三疣梭子蟹的幼体变态、个体形态大小和150日龄个体的生长特性产生的影响程度不同,造成一定程度的衰退。
关键词:三疣梭子蟹;自交;形态学指标;幼体变态;衰退
三疣梭子蟹Portunustrituberculatus隶属于甲壳纲Crustacea、十足目Decapoda、梭子蟹科Portunidae、梭子蟹属Portunus[1-3]。三疣梭子蟹分布于中国浙江、山东、广西、海南,以及朝鲜、日本、马来西亚等水域[4],因其具有肉鲜味美、营养价值与经济价值高等特点,20世纪80年代初以来被列为中国沿海渔业资源中重要的养殖对象[5]。近年来,随着中国海洋养殖产业的迅猛发展,养殖环境急剧恶化,病害频发,种质资源退化严重,造成了巨大的经济亏损,制约了水产业的可持续健康发展。自然捕捞环境中的梭子蟹无法满足市场需求,人工养殖该蟹成为缓解市场需求的主要途径,因此,研究三疣梭子蟹的苗种质量显得尤为重要。由于三疣梭子蟹亲本来源受到种质资源退化、水域捕捞数目有限等因素的影响,人工养殖亲本所占比例越来越大。因此,在苗种繁育过程中,样本不可避免地发生自交,从而导致后代优良性状衰退,且有日趋严重的现象,故探究亲本自交对后代产生的影响变得更为迫切[6]。
一般将有亲缘关系的雌雄个体发生的交配繁殖称为自交,而在群体上看,自交指某一性状上具有一定概率的同型交配[7]。自交能提高优秀基因纯合性,可以分离不同基因型的品系,提高蓄养种群样本的稳定性和均一性,还能使有害的隐性基因显露,成为剔除有害性状及检测种群质量的有利手段[6]。在人工养殖和天然环境条件下,均有很大概率引起自交,然而自交会使物种的体高、体长、颜色等一系列表型性状降低,称为自交衰退[7]。目前,有关水产动物鱼类、虾类、贝类自交衰退研究较多,主要集中于对样本的受精繁殖、生长发育、存活产量等相关生理性状方面[8-15],而对甲壳动物蟹类中相关的研究甚少,仅见高保全等[16]、王好锋等[17]对三疣梭子蟹在形态学及繁殖等方面的自交衰退情况有过报道。高保全等[16]研究发现,通过杂交能够改进三疣梭子蟹的生长发育,然而杂交具有局限性,其需要纯系的维持才能使之生效,并需要保持父母本的来源是不同群体,而同一群体若长期自交可能会导致生存能力和繁殖能力下降等问题;王好锋等[17]研究发现,六代家系近交的确降低了三疣梭子蟹的生长、存活和产量,相比于F1家系,近交系数每增加10%,就会引起全甲宽衰退-2.4%~-5.1%、体质量衰退-0.8%~-3.5%、存活率衰退-34.4%~-69.9%、产量衰退-14.1%~-35.4%。
本试验中,结合目前三疣梭子蟹自交的研究现状,以三疣梭子蟹自交家系为材料,从自交对三疣梭子蟹各个形态学指标产生的差异、生长、存活和产量等若干方面的影响进行深入研究,对人工养殖的三疣梭子蟹自交九代家系相关方面的数据进行分析,以期为三疣梭子蟹的选育工作提供参考。
1.1 材料
自2005年起,中国水产科学研究院黄海水产研究所在山东省潍坊市昌邑海丰水产养殖基地于室内通过人工交配的方式[17],建立了三疣梭子蟹自交家系,距今已传代至第九代(F9)。
1.2 方法
本研究中参照王好锋等[17]的试验方法。
1.2.1 试验亲蟹的指标测量 在三疣梭子蟹亲蟹受精并抱卵后,按期观测梭子蟹卵块颜色,用解剖镜查看卵内幼体成长情况,记录幼体每分钟心跳次数,用游标卡尺测量亲蟹形态学数据并标记,记录所测数据并据此估算亲蟹布池时间与排幼时间[18]。
1.2.2 幼体变态的测定 本研究中幼体变态将从排幼和变态率两个方面进行分析[19]。
(1)排幼量的测定。根据做好标记的亲蟹抱卵后卵块颜色及测量数据,在其排幼前一天用箱笼将亲蟹放入单独的育苗池内,亲蟹排幼后6h,于池内布置6个取样位置,用量杯量取400mL池水,依次查数每个取样位置所取池水中梭子蟹Z1幼体的个数并取其平均值,计算亲蟹排幼量,据育苗池水体的体积(底面积8m2×高1.5m)计算即可。
(2)变态率的测定。据文献报道,溞状幼体和大眼幼体阶段需要各蜕皮4次和1次[20]。从膜内孵出到成长为第一期幼蟹,需要在22~25℃的环境中培育15~18d。用解剖镜观测梭子蟹幼体发育情况,以发育阶段的时间,即每次幼体蜕皮变态6h为设置测量幼体时间点,同排幼量测定方法一样,同样布置6个取样位置,用量杯量取池水,在溞状幼体4个阶段(Z1~Z4),统算各个取样位置所取到池水中仔蟹数,取其平均值,并根据育苗池水体的体积(8m2×1.5m)计算各个阶段的变态率[20]。
1.2.3 各家系幼体的培育和养殖 在室外建立养殖池(60m×40m×1.5m),将养殖池均分为十二格,用10目筛网隔开。当培养的梭子蟹幼体变态为Ⅱ期仔蟹时,各个培育池分别捞出2000只转移至室外,各个小格内放入对应家系的仔蟹并记录标号。繁育苗种期间,所有小格内的培育条件保持一致。
1.2.4 形态学数据的测量 形态学指标主要包括全甲宽、甲宽、甲长、体高等数据。在各梭子蟹家系4个生长阶段(日龄为80、100、120、150d),用卡尺测量任意打捞的30个梭子蟹个体的形态学指标,测量结果精确至小数点后两位(mm),并测量梭子蟹体质量(g),天平精确至小数点后一位。在150日龄时统计各自交家系存活个体数,依据所得数据解析不同梭子蟹家系间的整齐度。
1.3 数据处理
1.3.1 幼体变态差别与形态性状差别分析 采用SPSS19.0统计分析软件整理所获得的每个三疣梭子蟹自交家系各幼体变态与形态学指标的数据,并对各性状的平均值、标准差和变异系数进行描述统计分析,利用单因素方差分析(One-WayANOVA)对不同时间阶段所有试验组进行显著性分析,将差别显著的试验组间进行多重比较(LSD)。
1.3.2 整齐度分析 整齐度分析与各个形态性状的变异系数密切相关[21-22]。变异系数是另一个权衡数据中各个观测值变异幅度的统计量,表现数据总体的离散程度大小。变异系数愈小,则离散程度愈小,整齐度就愈高,反之则整齐度愈低。变异系数(Coefficientofvariance,CV)计算公式[21]为
CV=指标标准差/指标平均数。
采用SPSS19.0软件进行统计,解析150日龄时每个世代梭子蟹的各个形态学指标的变异系数,并进行整齐度对比。
1.3.3 自交系数与衰退系数分析 自交的程度及其遗传效应可用自交系数F(Coefficientofinbreeding)表示[10]。自交系数F指一个个体因为自交而引起异质基因减低时,同质基因所占的概率[10]。根据自交系数的变化,可以估计自交对后代产生的影响。其数值越大表明个体的基因越纯合。参照Keys等[10]的方法计算自交系数(FX),计算公式为
其中:n1为父本到共同祖先的世代数;n2为母本到共同祖先的世代数;FA为共同祖先自己的自交系数。
各性状的衰退系数(Inbreedingdepressioncoefficient,IDC)通过以下公式计算[10]:
其中:![]() 分别为自交家系和F1代家系各个观测指标的平均值;FInbred和F分别为近交家系的自交系数和F1代家系的自交系数。
分别为自交家系和F1代家系各个观测指标的平均值;FInbred和F分别为近交家系的自交系数和F1代家系的自交系数。
2.1 自交三疣梭子蟹幼体的变态分析
2.1.1 排幼量 经统计得到三疣梭子蟹F1、F2、F3、F4、F5、F6、F7、F8、F9代的排幼量分别为56.50±22.89、51.84±16.90、50.04±23.25、48.77±13.03、45.82±18.45、54.03±16.09、46.93±15.82、48.38±14.79、43.80±14.08万粒,说明随同自交的递进,自交系数的递增,三疣梭子蟹实际的排幼量并无出现明显跌幅或涨幅的趋势,对其各代实际排幼量进行分析,差异性亦不明显(P>0.05),但值得注意的是,三疣梭子蟹自交程度较高的F9代家系的排幼量比其他各代低。
2.1.2 变态率 从表1可见:三疣梭子蟹各个自交家系在各阶段的幼体变态率总体呈下降趋势,且在自交程度高的F9代中尤为明显,F9代在各阶段幼体变态率分析中均处于劣势,然而分析后显示,各代相互之间,后一代与前一代相比,在4个阶段中的变态率并无显著性差异(P>0.05)。
2.2 自交各世代家系个体形态学参数
2.2.1 不同日龄时的形态学性状参数 各自交世代家系三疣梭子蟹日龄从80~150d时的全甲宽、甲宽、甲长、体高和体质量的差异分析结果如图1~图5所示。从图1可见:日龄为80、100、120、150d4个阶段中,F2代全甲宽皆为最大,F9代皆为最小;在80d时,除F9代全甲宽与F1~F4代全甲宽有显著性差异外(P<0.05),其他各代之间(上一代与下一代相比)并无显著性差异(P>0.05);而到100d时,各代间的差异开始逐渐显著,F4代已经与F1、F2代产生显著差异(P<0.05);在120d时,F2代长势很好,F2代虽与F1代有显著性差异(P<0.05),但在150d时,F2代与F1代的差异又回到不显著状态(P>0.05),F6、F8代与F7、F9代比较,虽然F6、F8代长势较好,然而却明显小于F1、F2两代三疣梭子蟹(P<0.05)。
表1 各代家系各个阶段的变态率
Tab.1 SurvivalrateateachstagefromzoeaⅠ(Z1)to megalopa(M)ineverygenerationofasamefamily%
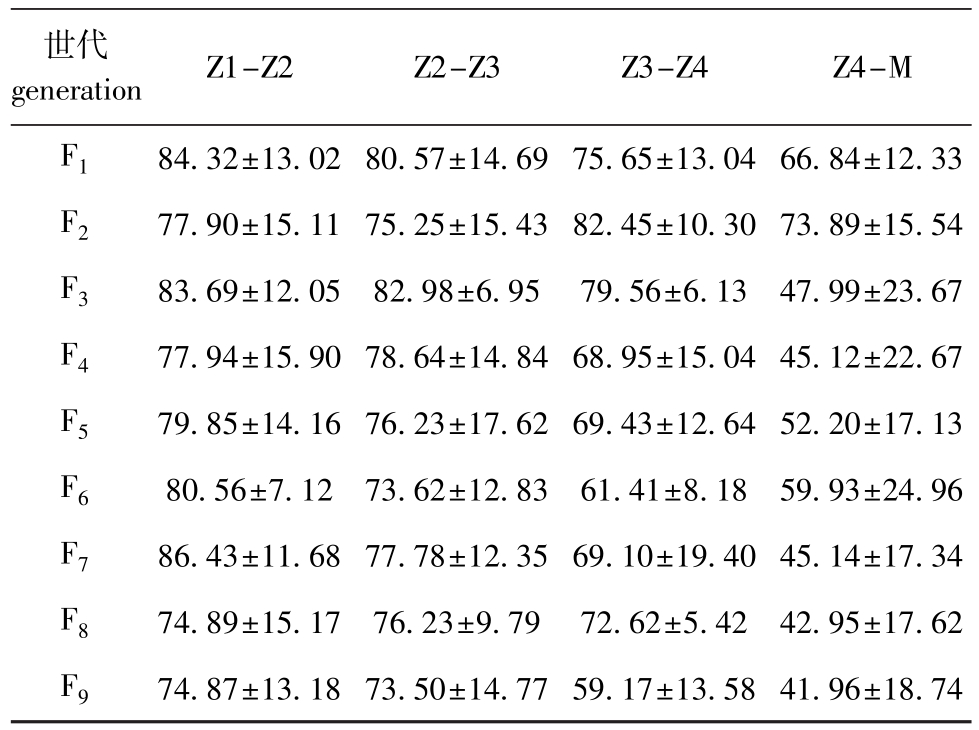
世代generation Z1-Z2Z2-Z3Z3-Z4Z4-M F184.32±13.0280.57±14.6975.65±13.0466.84±12.33 F277.90±15.1175.25±15.4382.45±10.3073.89±15.54 F383.69±12.0582.98±6.9579.56±6.1347.99±23.67 F477.94±15.9078.64±14.8468.95±15.0445.12±22.67 F579.85±14.1676.23±17.6269.43±12.6452.20±17.13 F680.56±7.1273.62±12.8361.41±8.1859.93±24.96 F786.43±11.6877.78±12.3569.10±19.4045.14±17.34 F874.89±15.1776.23±9.7972.62±5.4242.95±17.62 F974.87±13.1873.50±14.7759.17±13.5841.96±18.74
从图2可见:在80d时,三疣梭子蟹各代间甲宽均无显著性差异(P>0.05);在100~150d收获时,F2代的甲宽最大,F9代最小;F9代在100 d之后,与前三代的甲宽大小差距逐渐拉大,差异明显(P<0.05),而F4代之后各代间无显著性差异(P>0.05),但均呈下降趋势。
从图3可见:甲长总体情况与甲宽类似,在80d时,三疣梭子蟹各代间无显著性差异(P>0.05),但在100d时,甲长最小为F7代,在全甲宽与甲宽的分析中,F6代皆小于F3代,而在100d时,F6代的甲长略大于F3代,但无显著性差异(P>0.05),F6代这种生长优势在120、150d时却又渐渐削弱呈下降趋势。
从图4可见:80d时,随着三疣梭子蟹自交代数的增加,体高总体呈下降趋势,但各代间无显著性差异(P>0.05);虽然对各代间全甲宽、甲宽、甲长分析表明,在100d时已有显著性差异(P<0.05),但各代间体高在100d时无显著性差异(P>0.05),直到120d时,低自交家系才与高自交家系产生明显差异,而到150d时,随着自交代数增加体高呈下降趋势才逐渐明显。
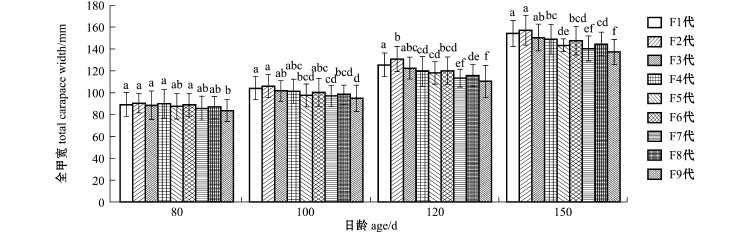
图1 三疣梭子蟹自交各世代家系全甲宽的比较分析
Fig.1 ComparativeanalysisoftotalcarapacewidthineachgenerationofswimmingcrabPortunustrituberculatus
注:同一日龄中标有不同小写字母者表示组间有显著性差异(P<0.05),标有相同小写字母者表示组间无显著性差异(P>0.05),下同Note:Themeanswithdifferentletterswithinthesameagearesignificantdifferencesatthe0.05probabilitylevel,andthemeanswiththesameletters withinthesameagearenotsignificantdifferences,etsequentia

图2 三疣梭子蟹自交各世代家系甲宽的比较分析
Fig.2 ComparativeanalysisofcarapacewidthineachgenerationofswimmingcrabPortunustrituberculatus

图3 三疣梭子蟹自交各世代家系甲长的比较分析
Fig.3 ComparativeanalysisofcarapacelengthineachgenerationofswimmingcrabPortunustrituberculatus
从图5可见:从80d开始到150d,F9代体质量便始终处于劣势,并与前面各代有显著性差异(P<0.05);到150d时,随着自交系数的增加各家系体质量呈明显下降趋势,各代间差异总体偏大。

图4 三疣梭子蟹自交各世代家系体高的比较分析
Fig.4 ComparativeanalysisofbodyheightineachgenerationofswimmingcrabPortunustrituberculatus
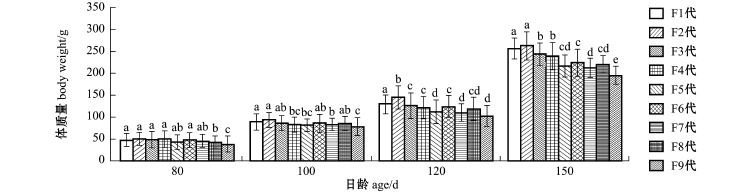
图5 三疣梭子蟹自交各世代家系体质量的比较分析
Fig.5 ComparativeanalysisofbodyweightineachgenerationofswimmingcrabPortunustrituberculatus
总体分析发现:在全甲宽、甲宽、甲长的比较分析中,三疣梭子蟹各个自交家系在80d时并无显著性差异(P>0.05),F1代无明显生长优势,而在100d之后的各阶段间,自交系数低的F1、F2代始终呈现出较优的生长状况,且优势明显,而从F3代开始便呈明显下降趋势,虽然F3代之后各代间普遍无显著性差异(P>0.05),但直到自交系数高的F9代时,一直处于生长劣势的F9代已经与低自交家系产生较大生长差异,F9代家系梭子蟹个头普遍偏小,衰退显著;而在体高分析中,三疣梭子蟹各自交家系生长在日龄为80、100、120、150d的4个阶段中虽然也呈同样下降趋势,但差异是于120d阶段时才表现出来;在体质量的分析中,与各项形态学生理指标不同,在80d时,三疣梭子蟹高自交家系F9代便表现出与低自交家系的体质量差异,且差异明显(P<0.05),直到150d收获时,三疣梭子蟹各代间的差异普遍明显,总体亦呈下降趋势;其中F6、F8两代高自交家系在各项指标分析中虽处于较好生长状态,且与低自交家系相比差异并不太明显,但总体上看,F6与F8代并没有偏离随着自交系数的增加,生长状况缓慢变差的这个趋势。
2.2.2 三疣梭子蟹各个自交家系群体150d龄时形态学性状参数的变异水平 如表2所示,150d时各自交家系各个形态性状的变异系数,随着自交代数的递增,全甲宽、甲宽、甲长、体高、体质量的离散程度皆呈先增大后减小的趋势,各个性状的变异系数变化趋势均几乎相同,其中F2代全甲宽、甲长、体高、体质量变异系数均较大、整齐度低,相较于其他家系自交程度极高的F9代5个性状的变异系数均较小,表明F9代在150d日龄阶段自交家系中,梭子蟹个体大小最整齐。
2.2.3 自交系数与衰退系数 本研究中所用样本是2005年采自舟山、莱州湾、海州和鸭绿江4个不同海域的野生三疣梭子蟹群体,理论上原始祖先的“祖父母本”在相隔距离上没有发生自交的可能性,所以初代三疣梭子蟹群体F1代的自交系数应为0。根据公式计算可得F1~F9代三疣梭子蟹每代家系的自交系数[10],结果如表3所示。
根据本研究中衰退系数的计算公式[10],以F1代家系作对比,计算可得自交引起的三疣梭子蟹各代样本各个性状的衰退系数。从表4可知,除F2代家系以外,随着自交系数的增加,相对于F1代,各性状的衰退情况,全甲宽为-6.12%~-21.95%,甲宽为-1.85%~-16.36%,甲长为-10.08%~-27.66%,体高为-9.39%~-22.70%,而体质量和排幼量的衰退系数相较于其他性状较高,体质量为-13.39%~-47.99%,排幼量为-9.03%~-45.13%。
表2 三疣梭子蟹各自交家系150日龄时形态学性状参数的变异系数
Tab.2 Variancecoefficientofmorphologicalparametersin 150daysoldswimmingcrabPortunustrituberculatusineachgeneration%
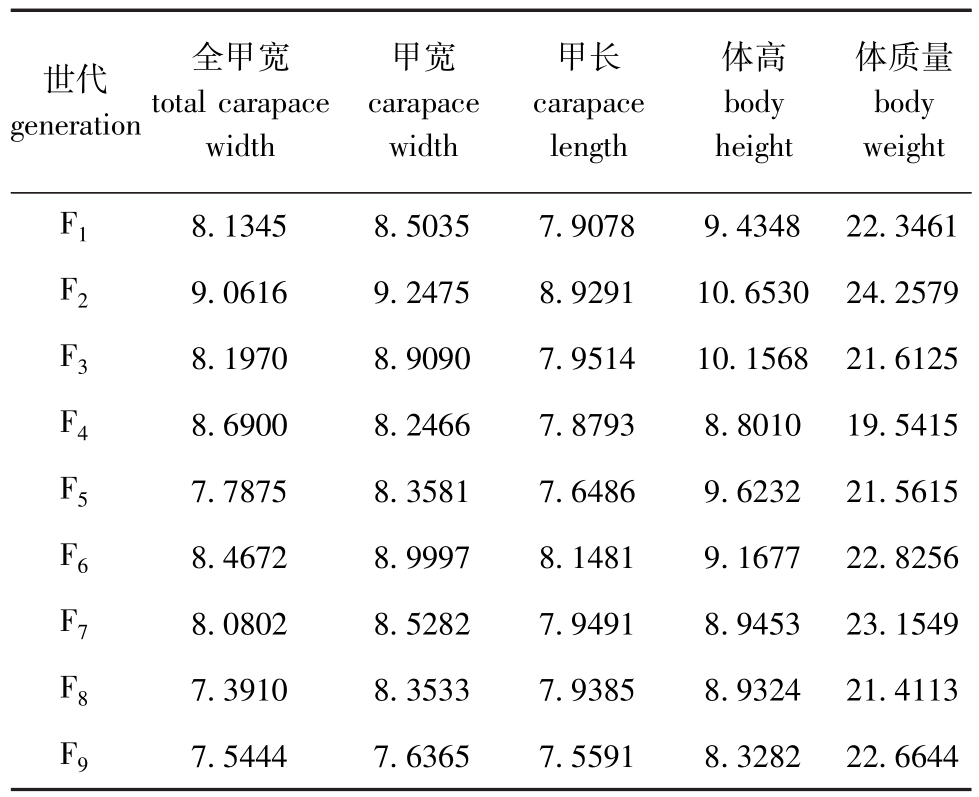
世代generation全甲宽totalcarapace width甲宽carapace width甲长carapace length体高body height体质量body weight F1 7.54447.63657.55918.328222.6644 8.13458.50357.90789.434822.3461 F29.06169.24758.929110.653024.2579 F38.19708.90907.951410.156821.6125 F48.69008.24667.87938.801019.5415 F57.78758.35817.64869.623221.5615 F68.46728.99978.14819.167722.8256 F78.08028.52827.94918.945323.1549 F87.39108.35337.93858.932421.4113 F9
表3 三疣梭子蟹自交各代亲蟹的自交系数
Tab.3 InbreedingcoefficientofswimmingcrabPortunustrituberculatusineachgeneration

世代generationF1F2F3F4F5F6F7F8F9自交系数inbreedingcoefficient00.25000.37500.43750.46880.48440.49220.49610.4980
表4 三疣梭子蟹近亲繁殖各代亲蟹的衰退系数
Tab.4 InbreedingdepressioncoefficientofswimmingcrabPortunustrituberculatusineachgeneration
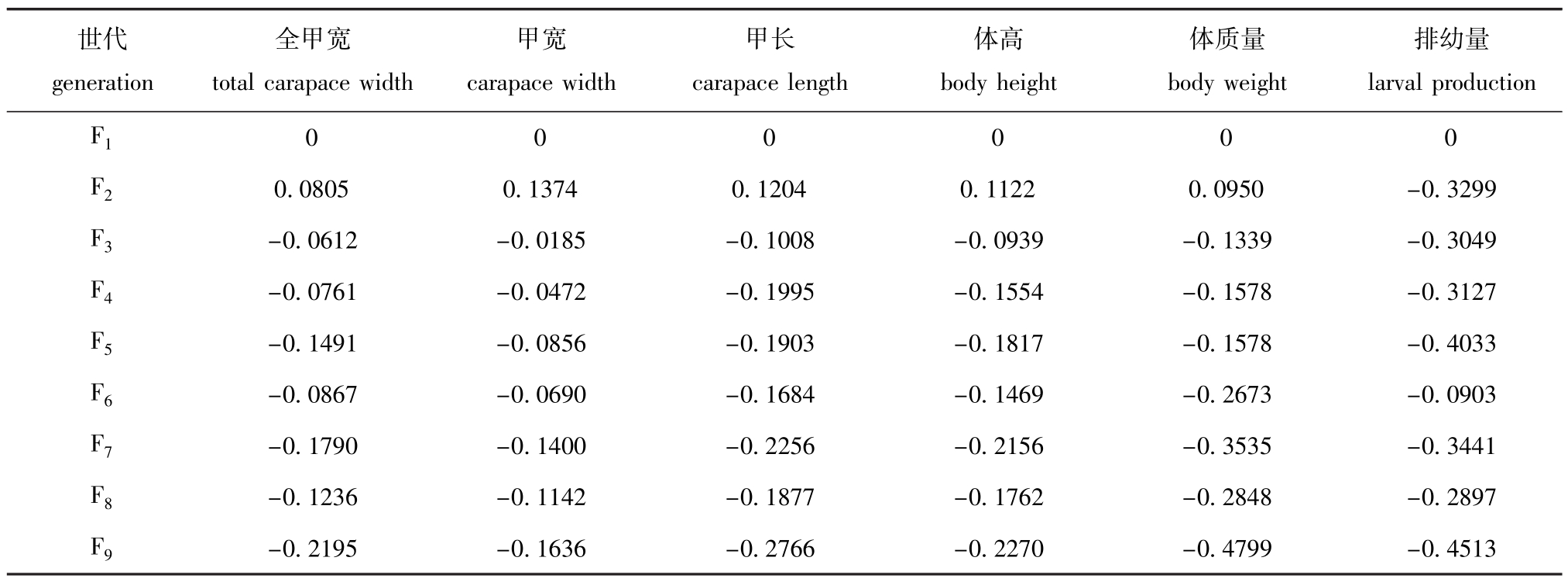
世代generation排幼量larvalproduction F1全甲宽totalcarapacewidth甲宽carapacewidth甲长carapacelength体高bodyheight体质量bodyweight 0 0 0 0 0 0 F20.08050.13740.12040.11220.0950-0.3299 F3-0.0612-0.0185-0.1008-0.0939-0.1339-0.3049 F4-0.0761-0.0472-0.1995-0.1554-0.1578-0.3127 F5-0.1491-0.0856-0.1903-0.1817-0.1578-0.4033 F6-0.0867-0.0690-0.1684-0.1469-0.2673-0.0903 F7-0.1790-0.1400-0.2256-0.2156-0.3535-0.3441 F8-0.1236-0.1142-0.1877-0.1762-0.2848-0.2897 F9-0.2195-0.1636-0.2766-0.2270-0.4799-0.4513
3.1 自交对衰退程度的影响
根据试验测量要求的不同、自交程度不一、物种的差别等,自交衰退的水平会有所差异,且衰退通常表现在生长性状(如成体体长、体宽、体质量等)和适应性相关性状方面(如幼体死亡率、性腺成熟系数、产卵数、孵化率、成体存活率等)[23]。自交程度的迥异对生长特性性状有不一样的影响,马大勇等[23]研究表明,全自交比迟缓自交导致的形态性状退化程度高达三倍左右。本试验中,所用自交家系属于近亲兄妹交尾引起的全同胞自交,各世代自交发生概率均高于10%,应当属于快速自交,从2005年至今,该三疣梭子蟹繁育群体自交已传至9代,自交水平很高,因此,随着自交系数递增而产生的衰退程度偏大。用于本研究的三疣梭子蟹样本均经过多年优良品种挑选培育而得,并使之保存繁衍。优秀世代三疣梭子蟹家系的良种选育也许是自交衰退对原始几代家系的作用不明显的原因之一,在幼体变态方面,自交让梭子蟹9代自交家系的幼体变态产生少量的自交衰退,经过解析,梭子蟹各代家系间差异不显著,但从所得数据分析,衰退趋势仍然存在,不能因为影响甚微就加以忽视。而在三疣梭子蟹个体生长方面,本研究结果表明,全甲宽、甲宽等规格大小的衰退程度较低,但体质量的衰退水平偏高。随着自交系数的增高,自交对三疣梭子蟹形态性状的影响较为明显,这也许会对将来的经济效益产生影响。5个测定指标的变异系数随着自交代数的递增,略呈愈来愈小的趋势,F9代所有变异系数几乎均呈最小,说明此世代的成体梭子蟹大小规格均衡程度稳定,整齐度最好,推测F1代属于最初源样本,属于野生三疣梭子蟹,未进行人为自交繁殖,来自不同区域的个体差异受不同的天然环境影响较大,所以个体大小不一;5个测量指标的变异系数较大,而随着人工繁育产生的高自交家系,在统一的养殖环境与条件下经过多年繁育,F9代个体形态指标受环境变换影响已经降低,笔者推测,可能在自交过程中,个体小的梭子蟹在群体进化的历史里已被自然选择所淘汰,群体的形态指标趋于平稳。也就是说,从总体上看,随着自交代数的增加,自交系数的增大,三疣梭子蟹的繁殖与生长各项指标均呈现出不同程度的自交衰退,且群体的个体规格大小也从F1代的大小不一到F9代的渐渐平稳。
3.2 自交“耐受”现象
在本试验中,三疣梭子蟹F1代并不是9代家系中所有指标最好的,而F2代的各项指标均接近最大,说明自交在一定程度内控制,会出现“耐受”现象,并在一定程度上抑制自交所带来的坏处,保留了优秀特性性状,留存并发展了优异个体样本的血缘。相比F3代,F6代与F8代在120龄以前也并没出现明显衰退,也可说明在个体生长过程中,高自交家系F6代与F8代在一定程度上表现出了耐受能力。呈现出这样自交衰退水平高低不同情况的缘由,也许是因为个体进化历史的差异,致使一些个体在进化历程里将有害的等位基因剔除,而在进化过程中另外有的个体可能发生了新型有害的突变,从而导致后代发生较大水平的自交衰退[24]。
3.3 避免自交衰退的措施
人工养殖繁育三疣梭子蟹时,群体内的自交在所难免,产生的自交衰退等影响也是日渐暴露的问题,对经济效益的影响是长期存在的。在三疣梭子蟹的育种过程中,梭子蟹的形态与幼体变态是测评水产养殖优劣的关键,所以研究自交衰退对生长繁殖的影响是不容忽视的工作。马大勇等[23]认为,人工培育种群内自交的弊端只有经过增加对后代有作用的亲本数才可以改善。在生产实践中,杂交也频繁用来改善养殖的种类,尤其人工繁殖能培养很多代生物的生产性能,但是从隐性有害基因的角度考虑,杂交优势只是自交衰退的补偿,主要是一种更正人工繁育种群自交衰退的手段[25]。本试验在通过对连续9代三疣梭子蟹自交群体幼体变态和形态学指标的分析后,发现随着自交世代增加,无论从幼体生长还是到变态为成体,自交对三疣梭子蟹均有不同的衰退迹象与影响。由此可知,自交理应于实际生产中有所需求时才可使用,而有目的、有计划、有选择地遗传育种,才是最大限度从遗传学上提高优良三疣梭子蟹经济性能,把利益推向持续发展道路的最佳方法。
参考文献:
[1] 戴爱云,冯钟琪,宋玉枝,等.三疣梭子蟹渔业生物学资源的初步调查[J].动物学杂志,1977,12(2):30-33.
[2] 戴爱云,杨思谅,宋玉枝,等.中国海洋蟹类[M].北京:海洋出版社,1986:213-214.
[3] 孙颖民,闫愚,孙进杰.三疣梭子蟹的幼体发育[J].水产学报,1984,8(3):219-226.
[4] HamasakiK,FukunagaK,KitadaS.BatchfecundityoftheswimmingcrabPortunustrituberculatus(Brachyura:Portunidae)[J]. Aquaculture,2006,253(1-4):359-365.
[5] 吴常文,虞顺成,吕永林.梭子蟹渔业技术[M].上海:上海科学技术出版社,1996:28-31.
[6] 张洪玉,罗坤,孔杰,等.近交对中国明对虾生长、存活及抗逆性的影响[J].中国水产科学,2009,16(5):744-750.
[7] FrankhamR,GilliganDM,MorrisD,etal.Inbreedingandextinction:effectsofpurging[J].ConservationGenetics,2001,2(3):279-284.
[8] 杨翠华,孔杰,王清印,等.控制环境养殖下近交对中国对虾早期体重和抗WSSV性状的影响[J].水产学报,2007,31(2):226-234.
[9] 刘振,闫喜武,霍忠明,等.蛤仔白斑马品系的家系选择及近交效应研究[J].大连海洋大学学报,2013,28(4):355-360.
[10] KeysSJ,CrocosPJ,BurridgeCY,etal.Comparativegrowthand survivalofinbredandoutbredPenaeus(marsupenaeus)japonicus,rearedundercontrolledenvironmentconditions:indicationsof inbreedingdepression[J].Aquaculture,2004,241(1-4):151-168.
[11] AulstadD,KittelsenA.Abnormalbodycurvaturesofrainbowtrout(Salmogairdneri)inbredfry[J].JournaloftheFisheriesResearchBoardofCanada,1971,28(12):1918-1920.
[12] 周海森,罗世滨,张伟杰,等.近交对中间球海胆受精率、孵化率和幼体发育的影响[J].大连海洋大学学报,2013,28(5):445-449.
[13] LongwellAC,StilesSS.GametecrossincompatibilityandinbreedinginthecommercialAmericanoyster,Crassostreavirginica Gmelin[J].Cytologic,1973,38(3):521-533.
[14] BeatieJH,HershbergerWK,ChewKK,etal.BreedingforResistancetoSummertimeMortalityinthePacificOyster(Crassostreagigas)[M].Washington:WashingtonSeaGrantRep,1978:780-813.
[15] Lannan JE.Broodstock management of Crassostrea gigas:I.genetic and environmental variation in survival in the larval rearing system[J].Aquculture,1980,21(4):323-336.
[16] 高保全,刘萍,李健,等.三疣梭子蟹(Portunus trituberculatus)不同地理种群内自繁和种群间杂交子一代生长性状的比较[J].海洋与湖沼,2008,39(3):291-296.
[17] 王好锋,刘萍,高保全,等.近交对三疣梭子蟹若干经济性状衰退的影响[J].中国水产科学,2013,20(6):1157-1165.
[18] 祝世军.三疣梭子蟹亲蟹培育及提高其受精卵孵化率的研究[J].河北渔业,2005(4):9-10.
[19] Wu Xugan,Liu Zhijun,Yao Guigui,et al.Relationship between the organogenesis of hepatopancreas and the yolk utilization during embryonic developmentof swimming crab,Portunus trituberculatus[J].Zoological Research,2009,30(4):449-456.
[20] 王好锋,高保全,段亚飞,等.三疣梭子蟹家系自交与杂交对繁殖和子代早期生长的影响[J].大连海洋大学学报,2014,(2):114-120.
[21] 王好锋.三疣梭子蟹自交与杂交家系繁殖与生长性状的比较分析[D].青岛:中国海洋大学,2013.
[22] Bensten H B,Olesen I.Designing aquaculturemass selection programs to avoid high inbreeding rates[J].Aquaculture,2002,204(3-4):349-359.
[23] 马大勇,胡红浪,孔杰.近交及其对水产养殖的影响[J].水产学报,2005,29(6):849-856.
[24] Ramsey M,Seed L,Vaughton G.Delayed selfing and low levels of inbreeding depression in Hibiscus trionum(Malvaceae)[J].Australian Journal of Botany,2003,51(3):275-281.
[25] 王好锋,韩晓琳,段亚飞,等.三疣梭子蟹自交与杂交家系子一代生长和存活的比较[J].渔业科学进展,2014,35(3):74-81.
Influence of inbreeding in 9 successive generations of a fam ily on metamorphosis and morphology of sw imm ing crab Portunus trituberculatus
Abstract:Nine generations of swimming crab Portunus trituberculatus within a same family(F1-F9)were systematically bred to reveal the influence of inbreeding on larval development and morphology in swimming crab via breeding and growth with ANOVA statistical analysis on one direction.The results showed that there was decline in survival rate(metamorphosis)at each stage from zoeaⅠ(Z1)tomegalopa(M)in each generation,74.87%from Z1 to Z2,73.50%from Z2 to Z3,59.17%from Z3 to Z4,and 41.96%from Z4 to M,with theminimal survival in F9,though no linear relationship between survival and each inbreeding generation from F1to F9.Themorphometrical valueswere shown to be declined from F3to F9,theminimal value in F9,significant from the other generations in the 80 days.However,the uniformity analysis of150 days old swimming crab revealed that the F9had variation coefficient of 7.636 5 in carapace width,7.559 1 in carapace length and 8.328 2 in body height,the least variation coefficient in all the generations.There were influences of various generations on larvalmetamorphosis,larval size and growth characteristics of swimming crab,showing inbreeding depression phenomenon.During the production season,compared with the F1inbreeding generation by increasing the inbreeding coefficient,except for F2,the estimated inbreeding depression coefficient was-6.12%--21.95%in overall carapace width,-1.85%--16.36%in carapace width,-10.08%--27.66%in carapace length,-9.39%--22.70%in body height,-13.39%--47.99%in body weight,and-9.03%--45.13%in larval yield,with higher depression coefficient in body weight and larval production than in other morphometrical values.The findings suggest that inbreeding should be avoided in crab breeding programs and that have a purpose in planned selective breeding.
Key words:Portunus trituberculatus;inbreeding;morphological index;larvalmetamorphosis;depression
中图分类号:S968.25
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2016.05.001
文章编号:2095-1388(2016)05-0469-08
收稿日期:2016-01-12
基金项目:国家自然科学基金资助项目(41576147);泰山领军人才工程高效生态农业创新类计划(LJNY2015002);青岛海洋科学与技术国家实验室鳌山科技创新计划项目(2015ASKJ02)