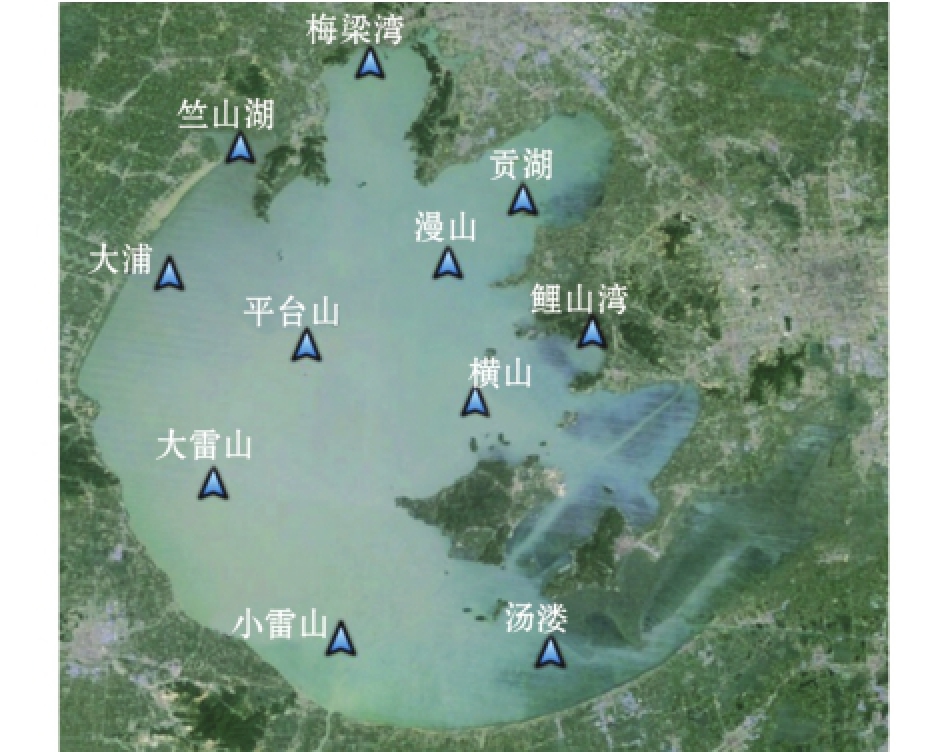
图1 采样站位示意图
Fig.1 Sketch of surveyed stations
摘要:为研究太湖麦穗鱼Pseudorasbora parva的种群结构、生长特征、死亡特征和资源利用状况,于2014 年1—12月使用虾笼网在太湖水域逐月进行了采样调查,并对采集的1207尾样本进行了分析。结果表明:麦穗鱼样本标准体长为27.03~107.36 mm,平均为 (60.03±13.29)mm,体质量为0.4~21.2 g,平均为(4.51±3.16)g;其体长与体质量呈幂函数相关,拟合关系式为W=2.1606×10-5L2.9537(R2=0.9323,n= 1207);Von Bertalanffy生长方程各参数为L∞=112.88 mm、K=0.46、t0=-0.51 a,体质量生长拐点为1.84 a;应用变换体长渔获曲线法估算总死亡系数 (Z)为1.87,利用Pauly经验公式估算自然死亡系数 (M)为1.07,捕捞死亡系数 (F)为0.80,开发率 (E)为0.43。研究表明,太湖水域麦穗鱼面临的捕捞压力较轻,处于适度利用状态,这与太湖水域无针对性捕捞且设有禁渔期有关,同时开捕期内肉食性鱼类面临的捕捞压力也有利于小型鱼类种群增殖。
关键词:太湖;麦穗鱼;体长;生长拐点;资源利用状况
麦穗鱼Pseudorasbora parva(Tamminck et Schlegel,1842)隶属于鲤形目Cypriniformes、鲤科Cyprinidae、鮈亚科 Gobioninae,原产地为东亚大陆,目前已扩散至除南极洲以外的全球范围[1],在中国的淡水水域广泛分布[2-7]。麦穗鱼栖息于水体的中下层,以浮游动物为主食,其生长较快、繁殖周期短、耐受力强[8-9],常用作行为[10-11]、毒理等试验的对象[12-15]。针对麦穗鱼的研究已有大量报道,主要集中于种群特征[16-18]、生理生化[19-21]和遗传结构[22-23]等方面,在中国西部高原湖泊,麦穗鱼的入侵机制也已受到关注[1,24]。
麦穗鱼作为一种小型、低值鱼类,其资源状况及捕捞状况往往不被重视。与此同时,在很多内陆水域特别是淡水湖泊中,麦穗鱼占据了较高的生态优势度,甚至会成为优势种[25-26]。从该角度考虑,作为鱼类群落结构的重要组成部分,其生长、死亡、利用强度等参数同样应予以关注。本研究中,以太湖麦穗鱼调查数据构建体长频率数据库,利用电子体长频率分析法ELEFANⅠ (Electronic length frequency analysis)[27]估算太湖麦穗鱼的生长和死亡参数,拟合其生长方程,并在此基础上评价了目前资源的利用状况,以期为麦穗鱼资源的评估与保护提供参考。
1.1 数据来源
基于均匀性和代表性原则[28]在太湖水域布设11个采样站位 (图1),使用虾笼网采集样本,网高和网宽均为30 cm,网目尺寸为0.8 cm,采样频率为每月1次。2014年1—12月共采集麦穗鱼样本1207尾。对采集的样本进行标准体长和体质量测定,分别精确至1 mm和0.1 g。按体长5 mm分组,用Excel 2010软件分析整理体长频率数据,利用国际水产生物资源管理中心 (ICLARM)与联合国粮农组织 (FAO)联合开发的体长基准鱼群评估系统数据分析软件FiSATⅡ,估算太湖麦穗鱼的生长与死亡参数,以及种群补充模式等参数。
1.2 估算方法
1.2.1 生长参数 麦穗鱼体长与体质量的关系使用幂函数方程描述:
图1 采样站位示意图
Fig.1 Sketch of surveyed stations
其中:W为样本的体质量;L为样本的体长;a和b为生长参数。
麦穗鱼的生长使用Von Bertalanffy生长方程描述:
建立麦穗鱼各月体长频率数据,采用ELEFANⅠ估算渐近体长L∞和生长参数K,应用Pauly经验公式[27]估算理论生长起点年龄t0,计算公式为
1.2.2 死亡参数 使用变换体长渔获曲线法估算总死亡系数 (Z)[29-30],计算公式为
其中:Nti为第i体长组中渔获个体数量;Δt为其从体长组下限生长到上限所需的时间;ti为第i体长组中值的年龄;a=ln N0,-b=Z,即总死亡系数的估算值。
借助 Pauly经验公式估算自然死亡系数(M)[27],计算公式为
其中:LT∞为渐近全长;K为生长参数;T为太湖水域年均水温。
根据调查数据拟合体长-全长关系式为
其中:LT为样本全长;L为样本体长,据此求出麦穗鱼的渐进全长LT∞。
2.1 渔获物结构
2014年1 —12月共采集到麦穗鱼样本 1207尾,体长范围为 27.03~107.36 mm,平均为(60.03±13.29)mm,其中体长介于41~70 mm的个体占据优势,所占比例为73.07% (图2);体质量范围为0.4~21.2 g,平均为 (4.51±3.16)g,其中体质量≤6 g的个体占据优势,所占比例为75.06% (图3)。
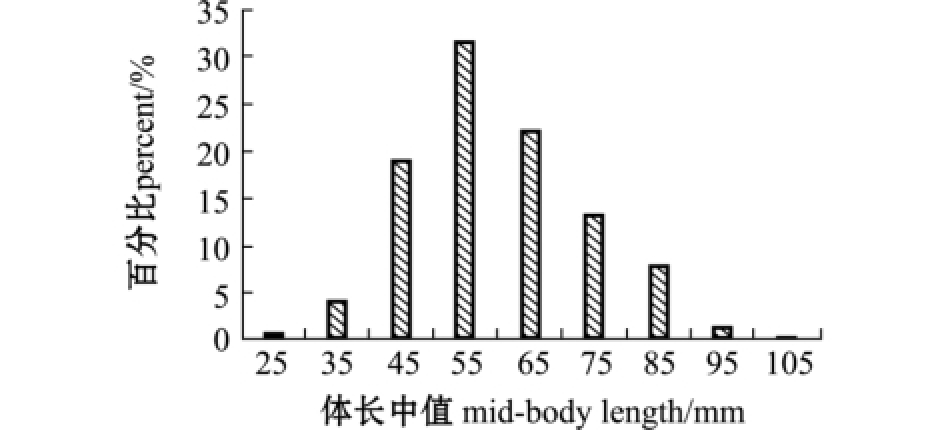
图2 麦穗鱼体长分布
Fig.2 Body length distribution of topmouth gudgeon
Pseudorasbora parva
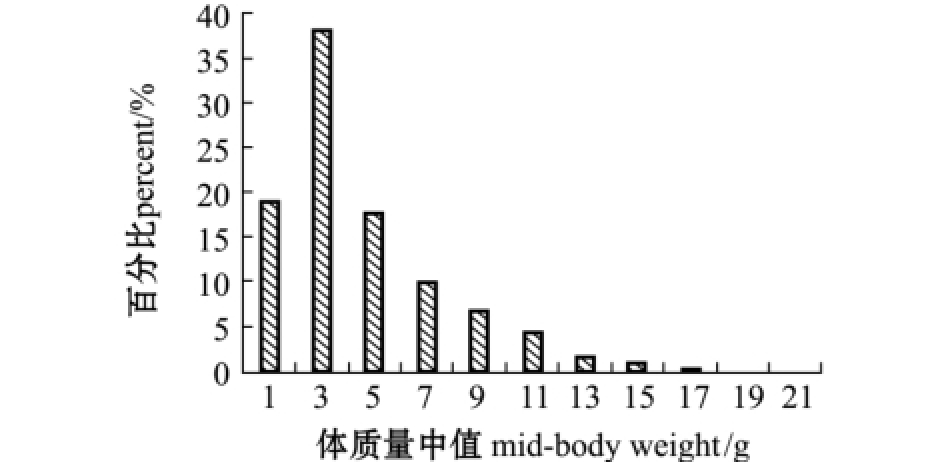
图3 麦穗鱼体质量分布
Fig.3 Body weight distribution of topmouth gudgeon Pseudorasbora parva
2.2 生长系数
2.2.1 体长频率及生长曲线 麦穗鱼体长 L (mm)与体质量W (g)呈幂函数相关 (图4),关系式为
1—12月麦穗鱼体长频率分布及生长曲线见图5,通过ELEFANⅠ拟合特殊的Von Bertalanffy生长方程,得到L∞=112.88 mm,K=0.46,生长方程为
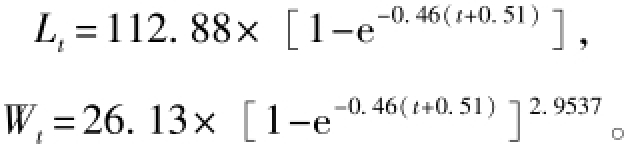
其中t0=-0.51 a,由公式 (3)计算得到,生长曲线拟合优度估计值ESP/ASP=0.246。
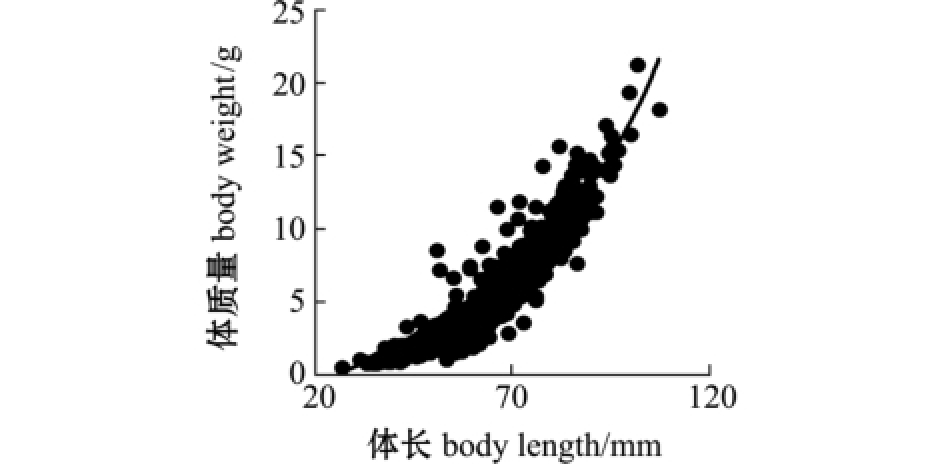
图4 麦穗鱼体长与体质量的关系
Fig.4 Relationship between body length and body weight of topmouth gudgeon Pseudorasbora parva
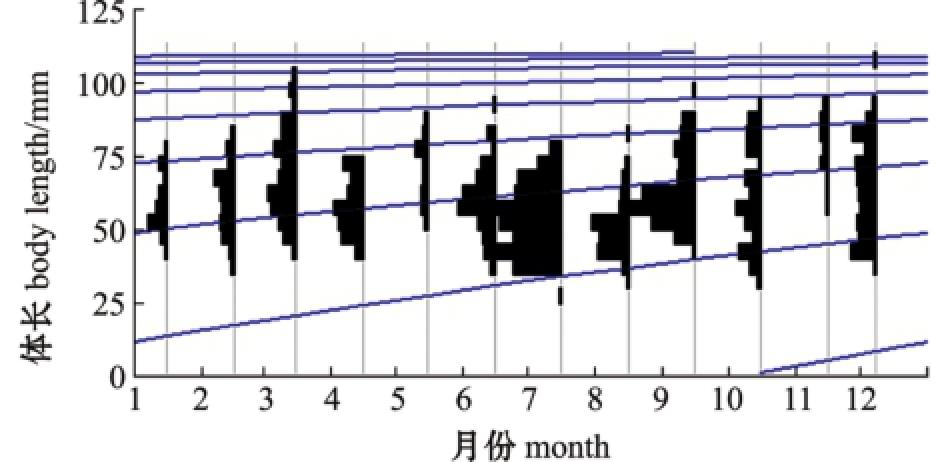
图5 麦穗鱼体长频率时间序列及应用ELEFANⅠ估计的生长曲线
Fig.5 Body length frequency and growth curve estimated by ELEFANⅠin topmouth gudgeon Pseudorasbora parva
2.2.2 生长速度及拐点 与多数鱼类相似,麦穗鱼体长为渐进生长,没有生长拐点,随着年龄的增加,体长趋近于渐进体长 (图6)。体质量的生长则表现为不规则的 “S”型曲线 (图7),拐点年龄ti=ln b/K+t0=1.84 a,对应体质量Wi=7.7 g。当t<1.84 a时,随着年龄的增大,麦穗鱼体质量生长速度上升,但生长加速度随之下降,体质量处于生长速度递增阶段;当1.84 a≤t<4 a时,随着年龄的增大,麦穗鱼体质量增长速度逐渐下降,生长加速度为负值,体质量处于生长速度递减阶段;当t≥4 a时,麦穗鱼体质量生长加速度到达最低点,下降趋势渐缓,体质量逐渐趋于渐进值,表明鱼体逐渐进入衰老阶段 (图8、图9)。
2.3 死亡系数
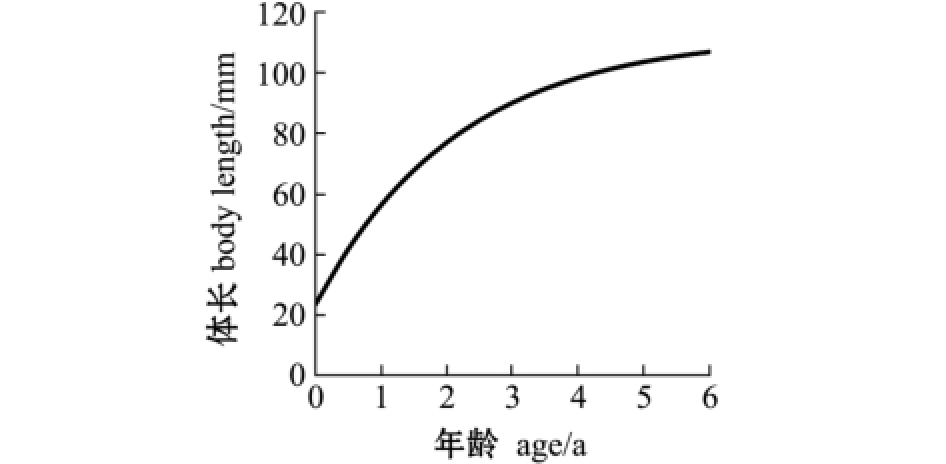
图6 麦穗鱼体长生长曲线
Fig.6 Body length growth curve in topmouth gudgeon Pseudorasbora parva

图7 麦穗鱼体质量生长曲线
Fig.7 Graphs of body weight increace in topmouth gudgeon Pseudorasbora parva
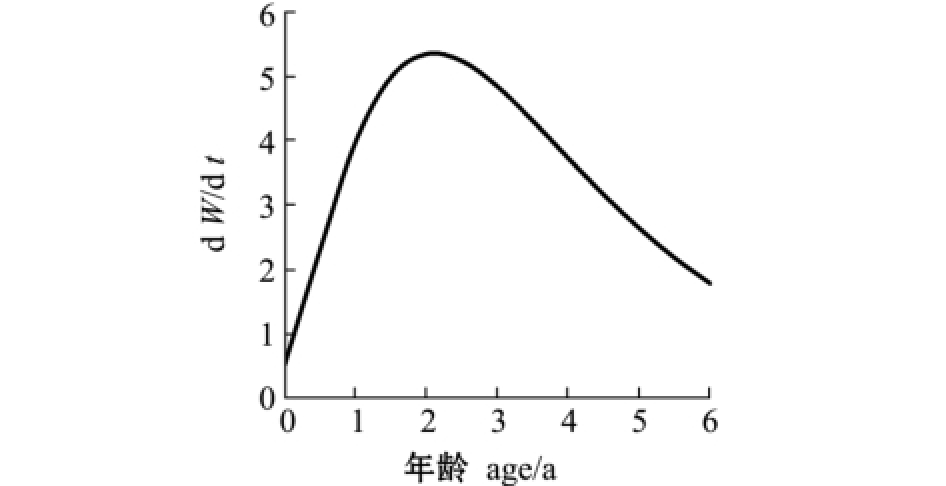
图8 麦穗鱼体质量生长速度曲线
Fig.8 Graphs of body weight growth rate in topmouth gudgeon Pseudorasbora parva
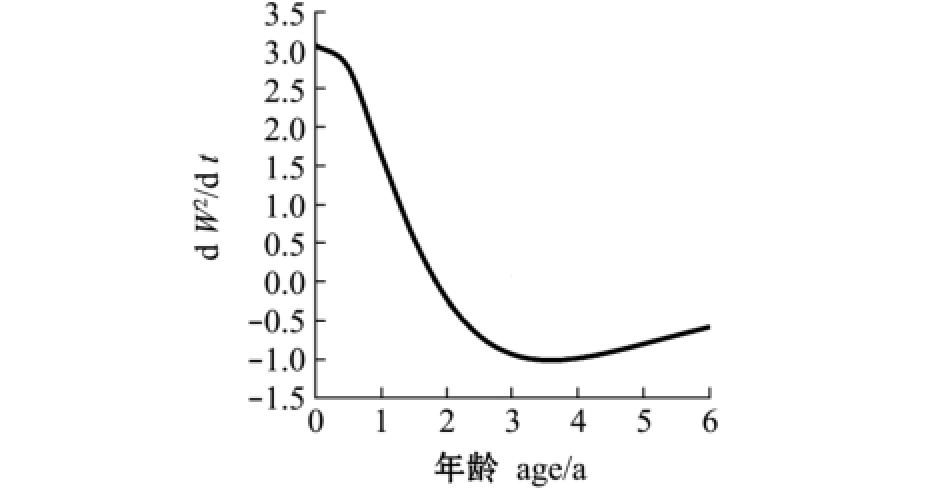
图9 麦穗鱼体质量生长加速度曲线
Fig.9 Graphs of body weight growth acceleration in topmouth gudgeon Pseudorasbora parva
2.3.1 总死亡系数 选取变换体长曲线中的8个点做线性回归 (图10),未达到完全补充年龄段最大体长组段 (最高点左侧)和体长接近L∞的年龄段的值不作为回归点,拟合的直线方程为
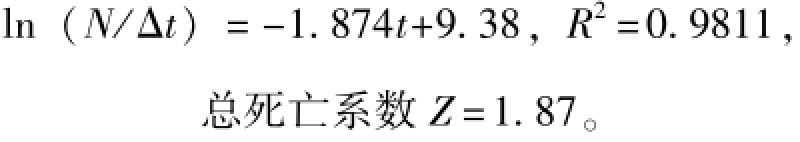
2.3.2 自然死亡系数 根据体长-全长直线方程推算渐进全长LT∞=136.26 mm;由LT∞=136.26 mm、K=0.46和太湖水域的全年平均水温17.3℃[31]根据公式 (5)计算出自然死亡系数 M= 1.07。
2.3.3 捕捞死亡系数 捕捞死亡系数F=Z-M= 1.87-1.07=0.80;开发率E=捕捞死亡系数F/总死亡系数Z=0.80/1.87=0.43。
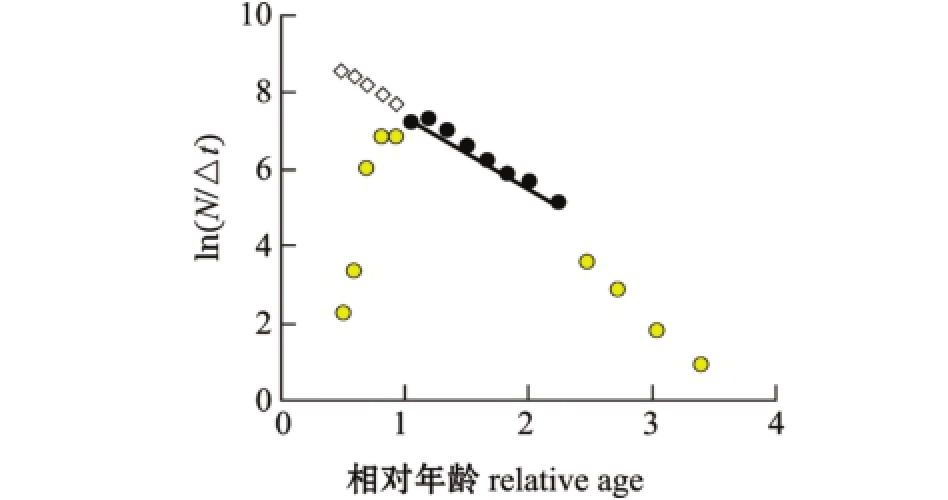
图10 总死亡系数估算
Fig.10 Estimation of mortality
2.4 资源利用状态评估
2.4.1 平均选择体长 根据变换体长渔获曲线保留百分数估计平均选择体长,由图10中拟合的线性关系,计算线性回归中未被使用的各点的ln(N/Δt)的观察值与期望值之比的累积率 (图11),当前渔获状态下,各个体长组麦穗鱼被捕获的概率随着体长的增加而增大,用Logistic曲线拟合,得出当麦穗鱼捕捞的平均选择体长Lc(保留达50%的体长)为46.04 mm,即开捕体长为46.04 mm。
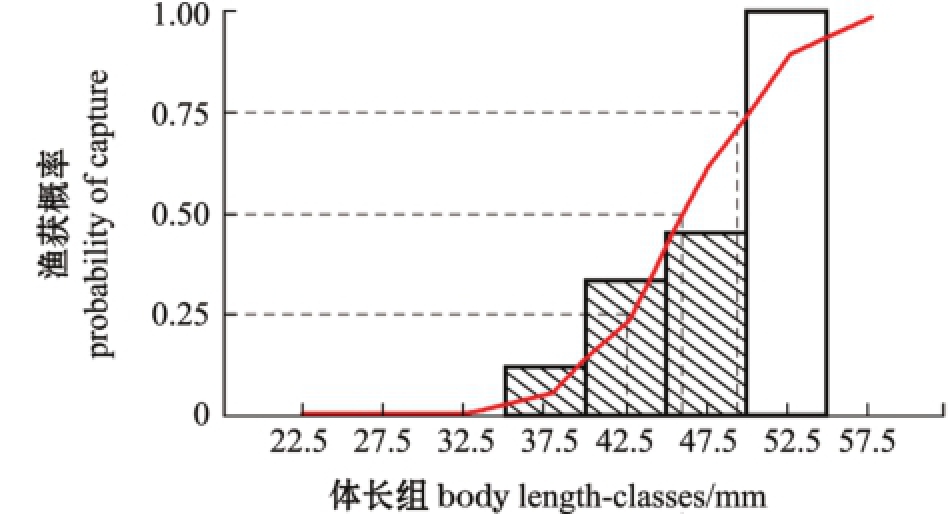
图11 渔获物变换体长曲线
Fig.11 Body length curve from converted catch
2.4.2 相对单位补充量 渔获量等值曲线图12中,P点为理想渔业点,M点为当前渔业点,Lc/L∞=0.408,开发率E=0.43,位于等值曲线的B象限,结果表明,太湖水域麦穗鱼面临的捕捞压力较小,处于适度利用状态,并未形成过度捕捞状态。
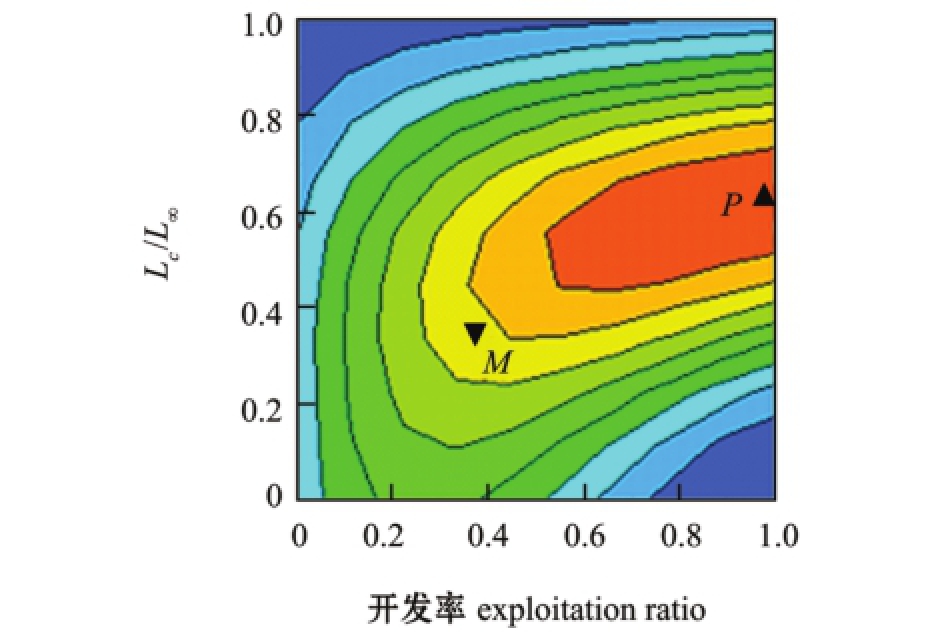
图12 单位补充量渔获量与开发率、开捕体长的关系
Fig.12 Relationship between relative yield-per-recruitment and exploitation ratio and body length at initial catch
本研究中,采集的麦穗鱼样本体长-体质量关系为 W=2.1606×10-5L2.9537,接近于等速生长[32-35]。样本优势体长组为27.03~107.36 mm,优势体质量组≤6 g。参照张堂林等[16]和杨竹舫等[36]的研究结果,本研究中采集的样本个体最大年龄应大于4龄,以1~2龄个体占据优势。麦穗鱼体长为渐进生长,没有生长拐点;体质量的生长拐点年龄为1.84 a,符合小型鱼类的生长特征[37]。生长参数K、总死亡系数Z、自然死亡系数M和捕捞死亡系数F分别为0.46、1.87、1.07和0.80,根据 M/K、e-K验证 M 值[38]、 K值[39]的合理性,结果表明,M/K=2.33,介于 1.5~2.5,e-K= 0.631<1,均符合条件。根据F和Z估算太湖麦穗鱼开发率E为0.43,表明太湖水域麦穗鱼种群面临的捕捞压力较轻,处于适度利用状态,这与已有相关研究报道的经济鱼类普遍处于过度利用状态[40-45]明显不同。
20世纪60—70年代,随着江湖联系的中断,加之湖泊围垦导致湖区鱼类产卵场和索饵场大量丧失,太湖水域逐步形成了以小型定居性鱼类占优的鱼类群落结构[46-48]。20世纪80年代初,太湖水域水质状况总体良好,以Ⅰ类和Ⅱ类水质为主,符合饮用水源地的水质要求[49]。20世纪90年代末至今,随着经济的快速发展,环太湖地区逐渐成为中国东部最发达的区域之一,与此伴随的是污染物大量排放、水域生态环境持续恶化。太湖水域呈现出明显的荒漠化趋势,鱼类的适宜栖息地急剧萎缩,加之捕捞强度未能得到有效控制,太湖水域渔业资源出现了显著的衰退迹象,具体表现为物种数量大幅减少、捕捞量急剧萎缩、主要捕捞对象趋于小型化和低龄化[6-7,50-51]。在此背景之下,鱼类群落中营养级较高的肉食性鱼类受到的影响更为显著,渔获比例大幅下降[52],这为小型鱼类种群的扩张提供了生态空间。
麦穗鱼作为一种分布范围广、适应性强的小型鱼类,并没有显著的经济价值。但从生态学角度而言,麦穗鱼在内陆淡水湖泊中有较大的天然资源量,通常具有较高的生态优势度[53],因此,其种群数量的稳定对于维持湖泊食物网结构的相对平衡具有生态学意义。此外,麦穗鱼主食藻类[18],对于控制湖泊营养负荷亦具有积极意义。太湖水域麦穗鱼并没有针对性的渔业捕捞,其主要的捕捞产出来自于以虾、蟹类经济物种为对象的虾笼网作业。目前太湖虾笼网1—4月没有捕捞作业,允许作业期为5—12月,部分避开了麦穗鱼的主要繁殖期(4—6月)[8-9],有助于麦穗鱼种群的补充及资源增殖。同时,太湖水域针对中、大型鱼类的捕捞压力并未显著缓解,由此留下的生态空间为以麦穗鱼为代表的小型鱼类种群扩张提供了有利条件,这也是目前太湖水域麦穗鱼种群利用率较低的重要原因。
参考文献:
[1] 张熙骜,隋晓云,吕植,等.基于Maxent的两种入侵性鱼类(麦穗鱼和鲫)的全球适生区预测[J].生物多样性,2014,22(2):182-188.
[2] 王金庆,宋祥甫,邹国燕,等.龙延河几种挺水植物上麦穗鱼卵的分布及其孵化率[J].淡水渔业,2014,44(1):46-52.
[3] 李晓雪,刘伟,唐富江,等.松花江哈尔滨段三种鮈亚科鱼类的食性与营养级分析[J].水产学杂志,2013,26(3):29-33.
[4] 路波,白艳勤,罗佳,等.葛洲坝下游中华鲟产卵场瓦氏黄颡鱼摄食活动的试验研究[J].生态学杂志,2013,32(10):2728-2734.
[5] 费骥慧,邵晓阳.高原湖泊鱼类生长特性与形态差异研究[J].海洋与湖沼,2012,43(4):789-796.
[6] 朱松泉.2002-2003年太湖鱼类学调查[J].湖泊科学,2004,16 (2):120-124.
[7] 朱松泉,刘正文,谷孝鸿.太湖鱼类区系变化和渔获物分析[J].湖泊科学,2007,19(6):664-669.
[8] 湖北省水生生物研究所鱼类研究室.长江鱼类[M].北京:科学出版社,1976:65-67.
[9] 倪勇,朱成德.太湖鱼类志[M].上海:上海科学技术出版社,2005:137-145.
[10] 王军,周应祺,钱卫国,等.实验室条件下麦穗鱼群体动态结构的表达[J].水产学报,2014,38(5):741-747.
[11] 王军.麦穗鱼群体结构与视觉关系的研究[D].上海:上海海洋大学,2014:1-63.
[12] 王晓南,刘征涛,闫振广,等.麦穗鱼物种敏感性评价[J].环境科学,2013,34(6):2329-2334.
[13] 张海艳,李霞,骆美蓉.苯酚和铬对麦穗鱼的急性毒性及苯酚对麦穗鱼的遗传毒性研究[J].安徽农业科学,2013,41(12):5389-5391.
[14] 顾晓军,李少南,樊德方.锐劲特与马拉硫磷混用对麦穗鱼的影响研究[J].农药学学报,1999,1(1):81-83.
[15] 徐晓丽,张彩军,吴剑峰,等.麦穗鱼对盐度和酸碱度的耐受性[J].湖北农业科学,2012,51(7):1423-1425.
[16] 张堂林,崔奕波,方榕乐,等.保安湖麦穗鱼种群生物学Ⅳ:种群动态[J].水生生物学报,2000,24(5):537-545.
[17] 张堂林,崔奕波,方榕乐,等.保安湖麦穗鱼种群生物学V:生产力[J].水生生物学报,2001,25(4):331-337.
[18] 杨瑞斌,边书京,周洁,等.梁子湖麦穗鱼食性的研究[J].华中农业大学学报,2004,23(3):331-334.
[19] 赵海鹏,黄林,张耀光.麦穗鱼的血液学特征[J].西南师范大学学报:自然科学版,2010,35(3):212-216.
[20] 梁拥军,孙向军,杨广,等.麦穗鱼营养成分组成测定与评价[J].安徽农业科学,2010,38(4):1869-1871,1973.
[21] 杨坤,祝东梅,王卫民.麦穗鱼耗氧率和窒息点的测定[J].水产科学,2013,32(5):256-260.
[22] 杨坤,王子健,祝东梅,等.麦穗鱼的组型分析和DNA含量测定[J].华中农业大学学报,2012,31(3):371-375.
[23] 马洪雨,郭金峰,岳永生.东平湖麦穗鱼群体遗传结构的微卫星标记分析[J].生命科学研究,2006,10(1):67-70.
[24] 曾燏.入侵种麦穗鱼的形态特征分析[J].水生态学杂志,2012,33(2):115-120.
[25] 叶少文,冯广朋,张彬,等.牛山湖小型鱼类群落结构特征及生物量估算[J].中国水产科学,2012,19(5):854-862.
[26] 叶少文,张堂林,李钟杰,等.牛山湖两种优势小型鱼类空间分布与沉水植被的关系[J].应用生态学报,2012,23(9):2566-2572.
[27] Pauly D.ELEFANⅡ:User's Instruction and Program Listings [M].Manila:ICLARM,1980.
[28] 张觉民,何志辉.内陆水域渔业自然资源调查手册[M].北京:农业出版社,1991.
[29] Moreau J,Cuende F X.On improving the resolution of the recruitment patterns of fishes[J].ICLARM Fishbyte,1991,9(1):45-46.
[30] Pauly D.Some simple methods for the assessment of tropical fish stocks[R].Makati,Metro Manila:FAO Fisheries Technical Paper,1983:52.
[31] 陈绍良,陆建伟,沈建强.太湖水体温度时空变化规律的初步研究[J].江苏水利,2009(3):38-39.
[32] Ricker W E.Computation and Interpretation of Biological Statistics of Fish Populations[M].New Jersey:The Blackburn Press,2000.
[33] 李文静,王剑伟,谢从新,等.厚颌鲂的年龄结构及生长特性[J].中国水产科学,2007,14(2):215-222.
[34] 田辉伍,岳兴建,陈大庆,等.怒江东方墨头鱼的年龄结构与生长特性[J].动物学杂志,2010,45(1):104-110.
[35] 刘飞,吴金明,王剑伟.高体近红鲌的生长与繁殖[J].水生生物学报,2011,35(4):586-595.
[36] 杨竹舫,李明德.天津地区麦穗鱼的生物学[J].动物学杂志,1989,24(1):11-14.
[37] 刘金殿,朱立新,卢伙胜,等.北部湾多鳞鱚生长与死亡参数的估算[J].浙江海洋学院学报:自然科学版,2010,29(1):64-69.
[38] Beverton R J H,Holt S J.A review of the life-spans and mortality rates of fish in nature,and their relation to growth and other physiological characteristics[J].CIBA Foundation Colloquia on Ageing,1959(5):142-180.
[39] 费鸿年,张诗全.水产资源学[M].北京:中国科学技术出版社,1990:303-305.
[40] 侯刚,朱立新,卢伙胜.北部湾二长棘鲷生长、死亡及其群体组成[J].广东海洋大学学报,2008,28(3):50-55.
[41] 陈作志,孔啸兰,徐姗楠,等.北部湾深水金线鱼种群参数的动态变化[J].水产学报,2012,36(4):584-591.
[42] 胡芬,严利平,李圣法.东海区刺鲳生长、死亡及资源利用状况评价[J].水产学报,2006,30(5):662-668.
[43] 孙远远,昝肖肖,徐宾铎,等.海州湾及邻近海域大泷六线鱼的生长、死亡和最适开捕体长研究[J].中国海洋大学学报:自然科学版,2014,44(9):46-52.
[44] 毕婷婷,侯刚,张胜宇,等.基于壳长频率分析的洪泽湖河蚬渔获物年龄结构、生长与死亡参数估计[J].水生生物学报,2014,38(4):797-800.
[45] 宋文,祝东梅,王艺舟,等.梁子湖团头鲂的年龄和生长特征[J].大连海洋大学学报,2014,29(1):11-16.
[46] 谷庆义,仇潜如.太湖鱼类区系的特点及其改造和调整的探讨[J].淡水渔业,1978(6):33-37.
[47] 邓思明,藏增嘉,詹鸿禧,等.太湖敞水区鱼类群落结构特征和分析[J].水产学报,1997,21(2):135-142.
[48] 李圣法,藏增嘉,邓思明,等.太湖敞水区鱼类种间关系现状[J].水产学报,1998,22(1):44-48.
[49] 刘雪芬.太湖水质污染状况的初步评价[J].海洋湖沼通报,1982(4):25-36.
[50] 唐晟凯,张彤晴,沈振华,等.太湖鱼类学调查及渔获物分析[J].江苏农业科学,2012(2):376-379.
[51] 唐渝.太湖湖鲚生长特征和临界年龄的研究[J].生态学杂志,1986,5(3):5-9,13.
[52] 刘恩生.太湖鱼类群落变化规律、机制及其对环境影响分析[J].水生态学杂志,2009,30(4):8-14.
[53] 张堂林.扁担塘鱼类生活史策略、营养特征及群落结构研究[D].武汉:中国科学院研究生院(水生生物研究所),2005:158-162.
Growth and mortality of topmouth gudgeon Pseudorasbora parva and evaluation on resource utilization in Taihu Lake
Abstract:Based on a total of 1207 samples of topmouth gudgeon Pseudorasbora parva collected by shrimp pot from January to December in 2014,an investigation was conducted monthly in Taihu Lake to study the composition,growth and mortality of capture as well as the resources utilization.The standard body length of samples ranged from 27.03 mm to 107.36 mm with mean value of(60.03±13.29)mm while the body weight ranged from 0.4 g to 21.2 g with the mean value of(4.51±3.16)g.A power function was presented between the body length and body weight while the relationship exhibited as follows:W=2.1606×10-5L2.9537(R2=0.9323,n=1207).The parameters of Von Bertalanffy growth equation were displayed as below:L∞=112.88 mm,K=0.46,t0=-0.51 a.The inflection point of body weight was about 1.84 years.Total mortality coefficient(Z)was 1.87 estimated by transformed body length catch curve method and natural mortality coefficient(M)was estimated about 1.07 with Pauly equation.Fishing mortality coefficient(F)was 0.80 and exploitation rate(E)was 0.43.The results showed that topmouth gudgeon confronting lower fishing pressure in Taihu Lake,stayed in a moderate utilization state which was related to non-targeted fishing and the enactment of forbidden fishing period in Taihu Lake,and the meanwhile,fishing pressure that carnivorous fishes faced in fishing period was beneficial to the proliferation of small-sized fish species as well.
Key words:Taihu Lake;Pseudorasbora parva;body length;inflection point;resources utilization condition
中图分类号:S931.1
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2016.04.004
文章编号:2095-1388(2016)04-0368-06
收稿日期:2015-10-11
基金项目:中央级公益性科研院所基本科研业务费专项 (2013JBFM08);江苏省自然科学基金资助项目 (BK2012093)