模拟海底水流和光照条件下光棘球海胆行为特征及其聚礁效果的初步研究
杨 军,刘永虎,田 涛,张国胜,陈 勇,尹增强,王 月,陈 辰
(大连海洋大学辽宁省海洋牧场工程技术研究中心,辽宁大连116023)
摘要:为观察光棘球海胆Strongylocentrotusnudus在饱食、饥饿和饥饿再投喂状态下的行为特征,及其对水泥、PVC两种材质模型礁的聚集特性,在实验室中模拟光棘球海胆栖息海域底层的昼夜水流速度和光照强度进行试验。结果表明:当光照强度达到560 lx、水流速度达到20 cm/s(9:00)时,均匀分布的光棘球海胆有90%向模型礁附近区域移动;当光照强度和水流速度下降时,光棘球海胆向模型礁附近区域外移动;运动趋势明显的光棘球海胆在饱食、饥饿和饥饿再投喂3种状态下的平均移动速度分别为0.039、0.041、0.052 cm/s,3种状态下光棘球海胆顺、逆水流运动次数的比值依次为饥饿状态 (1.63)>饱食状态 (1.18)>饥饿再投喂状态 (0.73),饥饿再投喂状态下光棘球海胆的活跃程度高于饱食和饥饿状态;鱼礁材质也影响光棘球海胆的聚集行为,光棘球海胆在饱食、饥饿和饥饿再投喂状态下对水泥模型礁的平均聚集率分别为25%、17%、24%,对PVC模型礁的平均聚集率分别为16%、7%、12%,光棘球海胆对两种模型礁的聚集效果均为饱食状态>饥饿再投喂状态>饥饿状态。研究表明,光棘球海胆在饱食和饥饿再投喂状态下对两种模型礁的聚集效果较好,而水泥材质的模型礁比PVC材质的模型礁聚集效果更好。
关键词:人工鱼礁;光棘球海胆;光照强度;水流速度;聚集率
光棘球海胆Strongylocentrotus nudues又名大连紫海胆,隶属于棘皮动物门、海胆纲、球海胆科,是食用海胆中重要的经济种类之一,主要分布于黄海、渤海部分海域的岛礁周围。20世纪80年代以来,由于过度采捕等原因使其野生资源已濒临枯竭[1]。实施海洋牧场建设,进行海胆的底播增殖是恢复海胆资源的有效手段,为此,辽宁、山东等省在沿海海域进行了大量的实践,并取得了良好的效果[2-3]。
目前,国内外研究人员对人工鱼礁功效及海珍品生物特性进行了大量研究,主要集中在人工鱼礁资源的调查及增养殖效果[4-10]、光照对刺参及模型礁对海珍品的行为影响[11-16]、不同材料和形状的增殖礁对海参聚集效果的影响[17-18]、大连紫海胆的生物学特性与食性[19-22]等方面,但光棘球海胆在不同光照强度和水流速度综合作用下的行为特征及人工鱼礁对其诱集效果的研究尚未见报道。为此,笔者在实验室水槽中,模拟光棘球海胆栖息海域的水流速度和光照强度,并放置两种材质的模型礁,研究在不同光照强度与水流速度下,不同材质的模型礁对光棘球海胆行为特征的影响以及对海胆的诱集效果,分析了人工鱼礁增殖区的适宜光照强度和水流速度,以期为光棘球海胆的科学底播及其养护与栖息场选址建设提供参考依据。
1 材料与方法
1.1 材料
试验用光棘球海胆采自大连市黑石礁海域,共100只,壳径为 (5.0±0.5)cm,壳高为 (3.0± 0.5)cm,体质量为 (80.0±0.5)g。在暂养池中暂养一周后,开始正式试验。暂养期间,每天投喂新鲜海带。暂养、试验用海水取自大连市黑石礁海域,经过滤沉淀后使用,水温为22.9℃,盐度为30,pH为7.84,DO为8.94 mg/L。
试验于2012年11月在辽宁省海洋牧场工程技术研究中心进行。
1.2 方法
1.2.1 光照强度和水流速度的设置 试验光照强度和水流速度的模拟值参照大连市黑石礁海域试验用光棘球海胆采捕区水深15 m海底处的连续观测值设定。
试验设定18:00—6:00的光照强度为0 lx,水流速度为5 cm/s,6:00—18:00的光照强度为0~560 lx,水流速度为0~30 cm/s(表1)。
试验光照采用60 W白炽灯8盏,15、25、40 W的白炽灯各4盏,垂直置于试验区域上方,实验室条件下经水下测试光照强度,将白炽灯分为4组,灯组的工作顺序如表1所示。
表1 试验参数
Tab.1 The parameters in the experiment
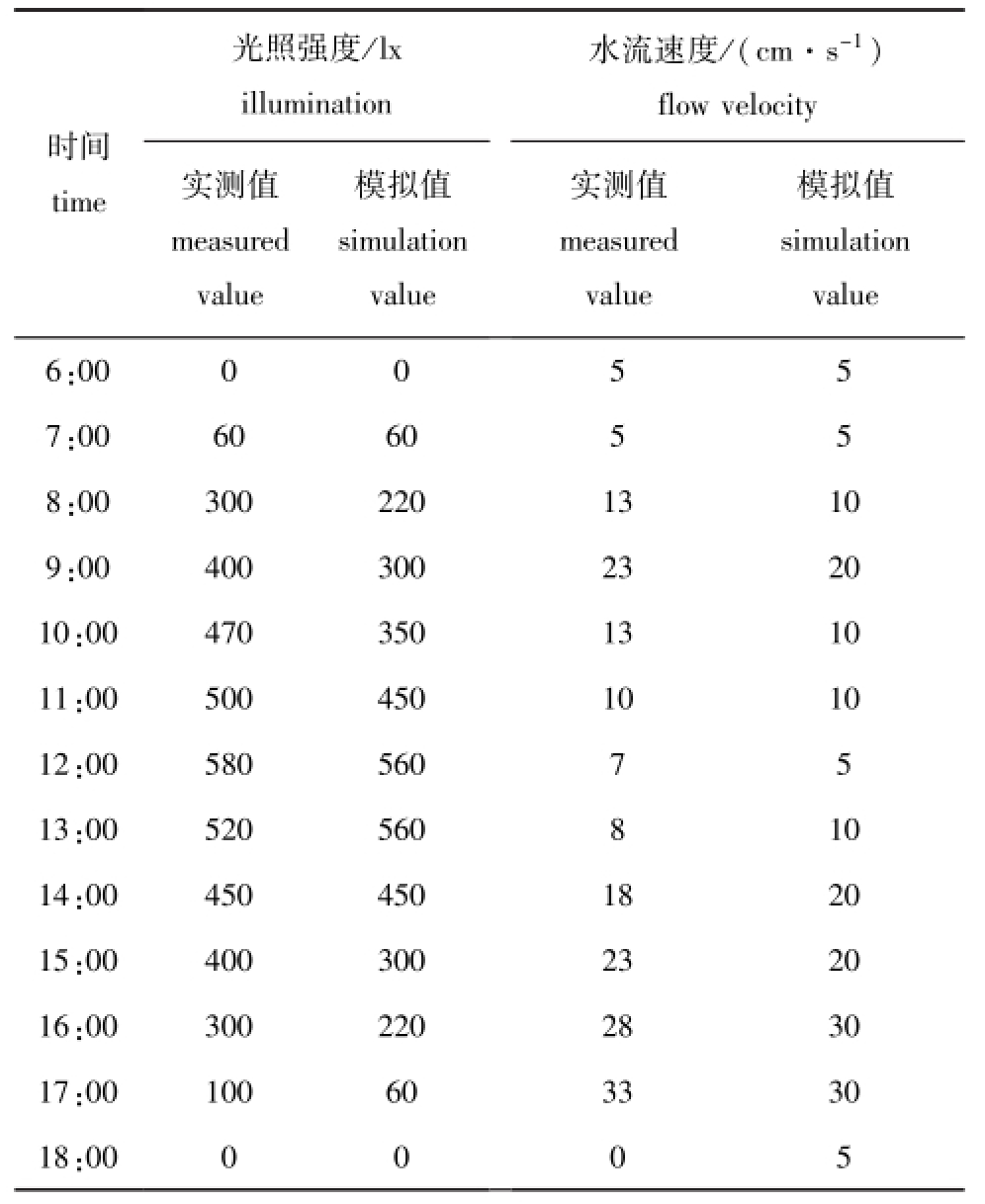
光照强度/lx 水流速度/(cm·s-1)时间time illumination实测值measured value模拟值simulation模拟值simulation flow velocity实测值measured value value 00 0 0 5 5 7:00 60 60 5 5 8:00 300 220 13 10 9:00 400 300 23 20 10:00 470 350 13 10 11:00 500 450 10 10 12:00 580 560 7 5 13:00 520 560 8 10 14:00 450 450 18 20 15:00 400 300 23 20 16:00 300 220 28 30 17:00 100 60 33 30 18:value 6:00 0 0 0 5
海胆行为特征试验在自制的环形水泥槽中进行,图1为试验区的构造示意图,其中环形水槽宽82 cm、长276 cm、水深100 cm。水流由简易造流机和整流板结合而成,流速变化由变频器改变电机转速完成。在试验区前部和后部焊接整流板,使水流趋于平稳,试验水槽内壁贴灰色PVC材料,以减小水槽内壁对水流的阻力,同时防止海胆的贴壁攀爬。
1.2.2 模型礁 模型礁选择水泥和PVC材料制作,用以对比不同材料礁体对海胆的诱集效果。水泥模型礁和PVC模型礁 (图2)的规格一致,均为30 cm×30 cm×30 cm,水泥模型礁孔窗为10 cm× 10 cm。将两种模型礁经海水浸泡15 d后,分别放置在试验区的中心位置。
1.2.3 试验方法
(1)区域划分。将试验区域划分为5个区域,即A区、B区、C区、D区和E区,每个区域长55.2 cm,其中B区是水泥模型礁诱集区域,D区为PVC模型礁诱集区域 (图1)。
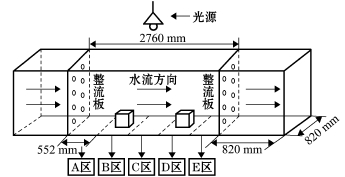
图1 水槽试验区示意图
Fig.1 The diagram of an experimental tank
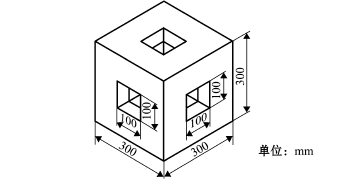
图2 模型礁
Fig.2 The artificial reef model
(2)聚集率试验。分别在饱食、饥饿和饥饿再投喂状态下进行海胆对水泥模型礁和PVC模型礁聚集率试验。分别选取30只饱食 (每天投喂)、饥饿 (饥饿6 d)和饥饿再投喂 (饥饿6 d投喂1 d)的光棘球海胆均匀放置在试验区域,连续3 d观察海胆的行为变化并记录海胆的聚集情况,试验期间不投饵。试验分3次进行,试验条件保持相同。海胆的行为特征观察试验与饱食状态下海胆对模型礁的聚集率试验同时进行。
试验过程实行全程观察监控 (鹰眼 KH-126HQ),其中监视器24 h连续观察,试验人员间隔1 h观察1次,并记录礁体附近光棘球海胆的数目。
1.2.4 指标的测定与计算 本研究中采用海胆运动总次数和移动速度来表征海胆在试验环境中的活跃程度。顺、逆水流运动次数比为海胆顺流和逆流运动的次数之比。移动速度定义为海胆在单位时间内的移动距离,其计算公式为
其中:v为移动速度 (cm/s);s为移动距离(cm);t为移动所用的时间 (s)。
采用平均聚集率作为分析指标,即在固定时间点观察到附着在礁体上的光棘球海胆数量的平均值占试验用海胆总数的百分比,其计算公式为
其中:P为平均聚集率 (%);X为观察期间对应各时间点附着在礁体上的海胆数量的平均值(只);N为该组试验海胆总数 (只)。
1.3 数据处理
采用Excel 2007软件和SPSS 22.0软件对试验数据进行处理,以分析不同环境条件下及不同喂食状态下海胆的行为特征。
2 结果与分析
2.1 不同光照强度和水流速度条件下光棘球海胆的行为特征
饱食状态下观察海胆在不同光照强度和水流速度条件下各时间点的行为特征。结果表明:在试验开始阶段 (18:00—6:00),当光照强度为0 lx、水流速度为5 cm/s时,B区平均有10只海胆,D区平均有5只海胆;当光照强度逐渐增强至220 lx、水流速度逐渐增强至10 cm/s(8:00)时,光棘球海胆开始向模型礁内部或附近移动;当光照强度增加至 300 lx、水流速度增加至 20 cm/s (9:00)时,移动到礁区的海胆数目达到最大,共有27只海胆在B区和D区;此后,受光照强度和水流速度双因素的影响,海胆在礁区的数目均未达到最大。
3种试验状态下,随机选取3只光棘球海胆对其移动次数和移动速度进行分析。结果表明:饱食状态下,海胆的运动活跃时间段为每日7:00—10:00,此时间段内海胆移动次数占海胆总移动次数的70.8%;饥饿状态下,海胆的运动活跃时间段为每日的13:00—16:00,此时间段内海胆的移动次数占海胆总移动次数的66.7%;饥饿再投喂状态下,海胆的运动活跃时间段为每日的13:00—16:00,此时间段内海胆的移动次数占海胆总移动次数的38.5%。这表明,饱食状态下海胆的运动活跃时间段集中在上午,饥饿和饥饿再投喂状态下海胆的运动活跃时间段集中在下午 (表2)。
饥饿状态下,3只光棘球海胆顺流和逆流运动次数比最大,为1.63,分别为饱食 (1.18)和饥饿再投喂状态下 (0.73)的1.38倍和2.23倍,而饥饿状态下海胆的逆流平均移动速度也最大,为0.074 cm/s,分别为饱食状态 (0.048 cm/s)和饥饿再投喂状态 (0.043 cm/s)的1.54倍和1.72倍(表2),这是由于光棘球海胆在饥饿状态下喜顺流而行,逆流行进时平均移动速度也较大,以便于快速找寻食物或避护场所。
饥饿再投喂状态下,3只光棘球海胆移动的总次数最多,为26次 (顺流11次,逆流15次),饥饿状态下,光棘球海胆移动的总次数最少,为21 次 (顺流13次,逆流8次),3种状态下光棘球海胆移动的总次数依次为饥饿再投喂状态>饱食状态>饥饿状态 (表2)。饥饿再投喂状态下,光棘球海胆的平均移动速度最大,为0.052 cm/s,饱食状态下,光棘球海胆的平均移动速度最小,为0.039 cm/s,3种状态下光棘球海胆的平均移动速度依次为饥饿再投喂状态>饥饿状态>饱食状态(表2)。根据海胆移动总次数和平均移动速度综合评价可得,饥饿再投喂状态下光棘球海胆的活跃程度明显高于饱食和饥饿状态。
表2 不同条件下光棘球海胆的行为特征
Tab.2 Behavior characteristics of the sea urchin Strongylocentrotus nudus under different conditions
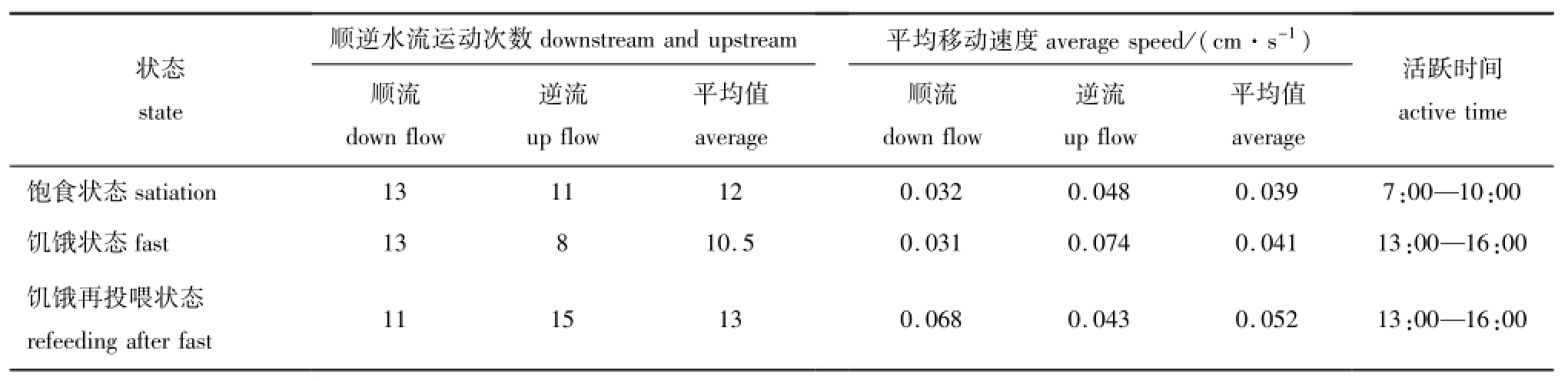
顺逆水流运动次数 平均移动速度average speed/(cm·s-1)状态state downstream and upstream顺流down flow活跃时间逆流up flow平均值 平均值active time average顺流down flow逆流up flow average饱食状态satiation 13 11 12 0.032 0.048 0.039 7:00—10:00饥饿状态fast 13 8 10.5 0.031 0.074 0.041 13:00—16:00饥饿再投喂状态refeeding after fast 11 15 13 0.068 0.043 0.052 13:00—16:00
2.2 不同光照强度和水流速度条件下光棘球海胆对模型礁的聚集率
3种试验状态下,光棘球海胆在不同时间点对水泥模型礁和PVC模型礁平均聚集率的变化如图3和图4所示。
光棘球海胆对水泥模型礁平均聚集率的变化为:饱食状态下,在8:00、11:00时平均聚集率最高,均为 66.67%;饥饿状态下,在8:00、9:00、10:00和12:00时平均聚集率最高,均为26.67%;饥饿再投喂状态下,在10:00时平均聚集率最高,为60% (图3)。同时,通过夜间摄像观察发现,0:00~1:00时光棘球海胆趋离水泥模型礁,分散在试验区域,但B区和D区分布居多。
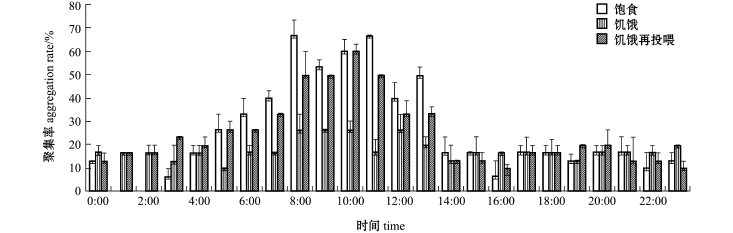
图3 光棘球海胆对水泥模型礁的平均聚集率
Fig.3 Average aggregation rates of the sea urchin in the concrete reef
光棘球海胆对PVC模型礁平均聚集率的变化为:饱食状态下,在12:00时平均聚集率最高,为60%;饥饿状态下,平均聚集率普遍低于20%;饥饿再投喂状态下,在12:00和13:00时聚集率最高,均为33.33% (图4)。这表明,光照强度和水流速度对光棘球海胆的聚集率影响较大,且白天的平均聚集率均高于夜间。当水流速度为10 cm/s、光照强度为350~450 lx时,3种状态的海胆对水泥模型礁的平均聚集率均较高。
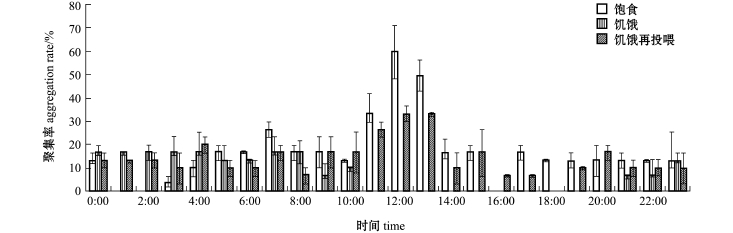
图4 光棘球海胆对PVC模型礁的平均聚集率
Fig.4 Average aggregation rates of the sea u rchin in the PVC reef
3种试验状态下,分别计算光棘球海胆对两种模型礁的平均聚集率。结果表明:饱食状态下,海胆对PVC模型礁的平均聚集率 (16%)低于水泥模型礁 (25%),夜间两种礁体的聚集效果差别不大;饥饿状态下,海胆对PVC模型礁的平均聚集率 (7%)也低于水泥模型礁 (17%),但夜间两种礁体的聚集效果差异明显,曾连续出现聚集率为0的现象;饥饿再投喂状态下,海胆的聚集情况与饱食状态类似,对 PVC模型礁的平均聚集率(12%)低于水泥模型礁 (24%),夜间两种礁体的聚集效果差别不大。这表明,无论哪种试验状态下,水泥模型礁的聚集效果明显高于PVC模型礁。
3 讨论
3.1 光棘球海胆的行为特性
本试验中模拟了自然海域底层的水流、光照和礁石环境。在自然条件下,海胆喜好附着于礁石上,昼伏夜出。试验暂养过程中,将海胆随机分布在暂养槽24 h后,大部分光棘球海胆白天附着在暂养槽壁上,出现这种现象与海胆的自然生活特性有关。陈勇等[17]通过研究模型礁对幼海胆行为的影响后认为,幼海胆喜欢在阴影较多且利于躲避敌害的地方栖息,具有避光性和避害性的生物特征,本试验结果与其研究结果一致。Kawamata[23]的研究表明,当水流速度超过3 cm/s时,海胆的摄食率显著下降,当水流速度超过4 cm/s时,海胆几乎停止摄食。流速增大会增加海胆抗击水流需要的能量,所以海胆会寻觅避水流区域。人工鱼礁多设置于水流较缓地区[24],海域中人工鱼礁区栖息的海胆数量往往高于其他海区,且水流速度也是影响海胆栖息的重要条件。
本次试验中,对处于饱食、饥饿和饥饿再投喂3种状态下的光棘球海胆,无论从海胆对模型礁聚集率的高低,还是从聚集率的稳定程度分析,饥饿状态下光棘球海胆的聚集率均远低于饱食状态和饥饿再投喂状态,因此,饵料效应是影响模型礁诱集效果的重要因素。通过对海胆行为的观察发现,无论饱食、饥饿还是饥饿再投喂状态,夜间海胆的聚集率均较低,这种现象可能与海胆夜间的觅食习性有关。
3.2 不同试验条件下光棘球海胆的活跃程度
在饱食状态下,光照强度和水流速度均会影响海胆的行为,在光照强度和水流速度逐渐增大的情况下,光棘球海胆向模型礁的内部或者附近移动。饱食状态下的海胆运动活跃时间多集中在上午,饥饿和饥饿再投喂状态下的海胆运动活跃时间多集中在下午。随机选取3只光棘球海胆的移动数据进行分析,结果表明,光棘球海胆顺、逆流总运动次数依次为饥饿再投喂状态>饱食状态>饥饿状态,平均移动速度依次为饥饿再投喂状态>饥饿状态>饱食状态。根据总移动次数和平均移动速度综合评价可得,海胆在饥饿再投喂状态下的活跃程度明显高于饥饿和饱食状态。
3.3 不同试验条件下光棘球海胆的聚礁效果
饱食状态下,光棘球海胆在11:00(水流速度10 cm/s、光照强度450 lx)时,对水泥模型礁的平均聚集率最高,为66.67%,在12:00(水流速度5 cm/s、光照强度560 lx)时,对PVC的平均聚集率最高,为60%;饥饿状态下,光棘球海胆在 8:00、12:00(水流速度5~10 cm/s、220~560 lx)时,对水泥模型礁的平均聚集率最高,均为26.67%,在7:00和8:00(水流速度5~10 cm/s、光照强度60~220 lx)时,对PVC模型礁的平均聚集率最高,为17.78%;饥饿再投喂状态下,光棘球海胆在10:00(水流速度 10 cm/s、光照强度350 lx)时,对水泥模型礁的平均聚集率最高,为60%,在12:00和13:00(水流速度5~10 cm/s、光照强度560 lx)时,对PVC模型礁的平均聚集率最高,为33.33%。这表明,在水流速度较小、光照强度较大 (11:00~13:00)的条件下,饱食状态的光棘球海胆对两种模型礁的平均聚集率均高于饥饿再投喂状态和饥饿状态,而饥饿状态下的光棘球海胆对两种模型礁的平均聚集率最差。这是因为饥饿状态迫使光棘球海胆适应不适环境寻找食物,而饱食和饥饿再投喂状态下,光棘球海胆更趋向于避光避流的水环境,这与陈勇等[17]关于模型礁对幼海胆行为影响的研究结论一致。
除水流速度和光照强度影响光棘球海胆的聚集率外,模型礁的不同材质也对光棘球海胆聚集率产生影响。光棘球海胆在饱食、饥饿和饥饿再投喂状态下对水泥模型礁的平均聚集率分别为25%、17%、24%,对PVC模型礁的平均聚集率分别为16%、7%、12%,无论在哪种状态下,海胆对水泥模型礁的平均聚集率均明显高于PVC模型礁。这是因为相对于表面光滑的PVC模型礁,光棘球海胆更倾向于在表面粗糙、亲水性更好的水泥模型礁上附着。实际复杂海况下,人工鱼礁对海胆具有较好的聚集效果。
建议人工鱼礁区选址建设应选择海底水流速度低于10 cm/s的海域,人工鱼礁的制作材料宜使用亲水性好的水泥材质。
参考文献:
[1] 郭文场,丁向清,尚建勋,等.中国海胆种质资源养殖与保护[J].特种经济动植物,2013(9):9-11.
[2] 田涛.人工鱼礁资源增殖生态学的研究[D].青岛:中国海洋大学,2007.
[3] 王波,李有乐,房慧,等.虾夷马粪海胆生物学及增殖养殖技术[J].齐鲁渔业,1999,16(3):14-16.
[4] 李永刚,汪振华,章守宇.嵊泗人工鱼礁海区生态系统能量流动模型初探[J].海洋渔业,2007,29(3):226-234.
[5] 张虎,朱孔文,汤建华.海州湾人工鱼礁养护资源效果初探[J].海洋渔业,2005,27(1):38-43.
[6] 张虎,刘培廷,汤建华,等.海州湾人工鱼礁大型底栖生物调查[J].海洋渔业,2008,30(2):97-104.
[7] 张伟,李纯厚,贾晓平,等.人工鱼礁附着生物影响因素研究进展[J].南方水产,2008,4(1):64-68.
[8] 王诗关,魏绍善,谭忠云,等.在贝藻养殖区投放人工礁增殖刺参的研究[J].齐鲁渔业,1989(3):73-75.
[9] Blaxter JH,Sand Batty R S.The reactive perceptive field of the larval plaice:a three-dimensional analysis of visual feeding[J]. ICESSymp,1988,11:474.
[10] D'Anna G,Giacalone V M,Badalamenti F,et al.Releasing of hatchery-reared juveniles of the white seabream Diplodus sargetus(L.,1758)in the Gulf of Castellammare artificial reef area (NW Sicily)[J].Aquaculture,2004,233(1-4):251-268.
[11] 张硕,孙满昌,陈勇.人工鱼礁模型对大泷六线鱼和许氏平鲉幼鱼个体的诱集效果[J].大连水产学院学报,2008,23(1):13-19.
[12] 武模戈.光照对水生动物的影响[J].河南教育学院学报:自然科学版,2001,10(2):38-39.
[13] 陈勇,高峰,刘国山,等.温度、盐度和光照周期对刺参生长及行为的影响[J].水产学报,2007,31(5):687-691.
[14] 游奎,刘鹰,杨红生,等.不同光源及光照时间对凡纳滨对虾生长的影响[J].海洋科学,2005,29(5):1-5.
[15] 周显青,牛翠娟,李庆芬.光照对水生动物行为的影响[J].动物学杂志,1999,34(2):45-48.
[16] 周显青,牛翠娟,李庆芬.光照对水生动物摄食、生长和存活的影响[J].水生生物学报,2000,24(2):178-181.
[17] 陈勇,吴晓郁,邵丽萍,等.模型礁对幼鲍、幼海胆行为的影响[J].大连水产学院学报,2006,21(4):361-365.
[18] 崔勇,关长涛,万荣,等.海珍品人工增殖礁模型对刺参聚集效果影响的研究[J].渔业科学进展,2010,31(2):109-113.
[19] 常亚青,丁君,宋坚,等.海参、海胆生物学研究与养殖[M].北京:海洋出版社,2004.
[20] 王波.大连紫海胆的生物学特性及增养殖[J].齐鲁渔业,1996,13(1):25-27.
[21] 高绪生,孙勉英,李国友,等.大连紫海胆食性的初步探讨[J].水产学报,1990,14(3):227-232.
[22] 尤凯,曾晓起,陈大刚,等.青岛近岸海域马粪海胆摄食的实验生态学研究[J].生态学报,2004,24(5):1006-1014.
[23] Kawamata S.Effect of wave-induced oscillatory flow on grazing by a subtidal sea urchin Strongylocentrotus nudus(A.Agassiz)[J].Journal of Experimental Marine Biology and Ecology,1998,224(1):31-48.
[24] 杨吝,刘同渝,黄汝堪.中国人工鱼礁的理论与实践[M].广州:广东科学技术出版社,2005.
Behavior and aggregation of sea urchin Strongylocentrotus nudus to reefs under the simulated water current and illum ination at seafloor
YANG Jun,LIU Yong-hu,TIAN Tao,ZHANG Guo-sheng,CHEN Yong,YIN Zeng-qiang,WANG Yue,CHEN Chen
(Center for Marine Ranching Engineering Science Research of Liaoning,Dalian Ocean University,Dalian 116023,China)
Abstract:Behavior and aggregation of satiated,fasted and refed sea urchin Strongylocentrotus nudus were investigated in reefs prepared by two types of PVC under simulated water current and illumination at seafloor.The results showed that 90%of sea urchin evenly distributed was found to move near to the reef zone at light intensity of 560 lx,and flow velocity of 20 cm/s(9:00),while theymoved outoff the reefs at the declining light intensity and water velocity.The sea urchin hadmovement speed of0.052 cm/s,0.039 cm/s and 0.041 cm/swhen theywere satiated,fasted and refed,respectively,with the movement frequency ratio of along water to againstwater in the three states was described as:fasted(1.63)>satiation(1.18)>refed after fasted(0.73),indicating that refeeding after fasted led to higher activity than at satiation and fast.Also,the reefmaterial influenced the aggregation behavior of sea urchin.An average aggregation rate of25%,17%and 24%to the cement reefswas observed in the sea urchin which was satiated,fasted and refed,respectively,and there was average aggregation rate of 16%,7%and 12% to the PVC reefs in the sea urchin which was satiated,fasted and refed,respectively,showing the descendantorder of aggregation rate as satiation>feeding after fast>fast.The findings indicate that better aggregation rate was found in the satiated and refed sea urchin,and that there was better aggregation rate in the cement reefs than that in the PVC reefs.
Key words:artifical reef;Strongylocentrotus nudues;light intensity;flow velocity;aggregation rate
中图分类号:S953.1
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2016.02.018
文章编号:2095-1388(2016)02-0219-06
收稿日期:2015-07-06
基金项目:国家 “十二五”科技支撑计划项目 (2012BAD18B03,2013BAD23B02)
作者简介:杨军 (1985—),男,助理实验师。E-mail:yangj@dlou.edu.cn
通信作者:刘永虎 (1984—),男,实验师。E-mail:tiger@dlou.edu.cn