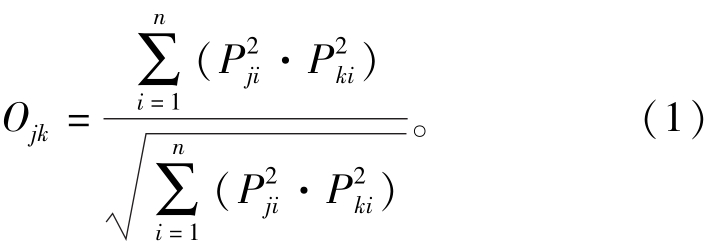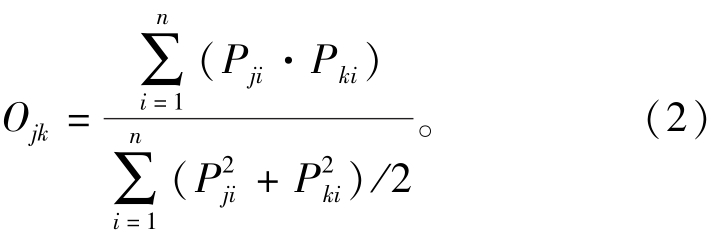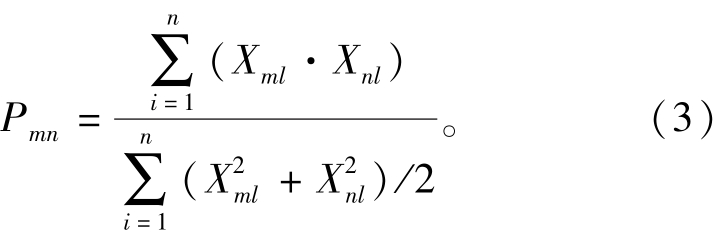基于栖息地适宜度指数模型的刺参环境适宜性初步研究
李林晗,陈勇,田涛,刘永虎,于亚群,张皓宇
(大连海洋大学辽宁省海洋牧场工程技术研究中心,辽宁大连116023)
摘要:为了评价刺参Apostichopus japonicus生存环境的适宜度,基于动物栖息地适宜度指数 (Habitat suitability index,HSI)模型,对刺参的环境适宜性进行了研究。选取温度、盐度、溶解氧、底质环境类型和底质砂砾含量作为影响刺参适应性的环境因子,根据经验赋值法和数理统计方法,绘制了大长山海域刺参各影响因子的适宜性指数 (Suitability index,SI)曲线图,并利用几何平均法综合估算HSI值。结果表明:2013—2014年7个航次调查的HSI平均值分别为0.66、0.59、0.67、0.46、0.66、0.65、0.65,冬季的适宜度 (HSI=0.46)明显低于其他季节,而春季 (HSI=0.66)及秋季 (HSI=0.67,HSI=0.65)的适宜度相对较高;同一航次中 (以2014年11月为例),7个站位的 HSI值分别为0.66、0.67、0.65、0.62、0.63、0.62、0.67,其中DCS01、DCS02和DCS07站位的适宜度较高。研究表明,春季及秋季在DCS01、DCS02和DCS07站位进行刺参苗种底播,刺参的成活率和生长情况可能更好。
关键词:刺参;栖息地适宜度指数;环境适宜性
刺参Apostichopus japonicus隶属棘皮动物门E-chinodermata、海参纲Holothuroidea、楯手目Aspidochirotida、刺参科Stichopodidae、仿刺参属Stichopus,是中国20多种食用海参中质量最好的一种,其营养价值和药用价值较高,被列为 “海味八珍”之一,并具有较高的经济价值[1]。刺参主要分布于中国辽东半岛、河北秦皇岛、胶东半岛等北方沿海区域。目前,商品类刺参生产主要有池塘养殖和底播增殖两种方式,且在自然海区底播增殖的刺参品质更佳。国内关于刺参增养殖的研究多集中在池塘养殖、近海围网养殖技术,以及与其他贝类的混养技术等,而对于其在环境适宜程度方面的研究较少。因此,探索一种综合的生境评价模型,有助于选择适合刺参底播的区域,可为实际增殖生产提供帮助。
栖息地适宜度指数 (Habitat suitability index,HSI)模型是一种评价野生生物生境适宜度程度的指数,最早被用来描述野生动物的栖息地质量[2]。HSI模型具有多种形式,一般表达式为 HSI=(SI1×SI2×SI3×…×SIn)1/n,其中SI为生境因子的适宜度指数,n为变量个数。目前,在渔业科学中常用的HSI综合算法有连乘法、最小值法、最大值法、几何平均法和混合算法等[3]。国外已开发出大量针对内陆、河流、湖泊和海洋渔业中的动物HSI模型,主要用于水生生物的保护与管理。Vinagre等[4]结合地理信息系统研究了葡萄牙塔古斯河口的欧洲鳎 Solea solea及塞内加尔鳎 Solea senegalensis的栖息地指数模型,并以此推测出两个适宜度较高的地区。Gore等[5]研究了小龙虾 Orconectes neglectus的适宜栖息地时空变化,结果表明,小龙虾不同生活史阶段的适宜栖息地不同。Soniat等[6]利用6~8个影响因子建立了墨西哥湾美洲牡蛎Crassostrea virginica的生境适宜度指数模型,并利用回归模型对其进行了改进。
国内也有较多关于动物HSI建模的报道。朱丽娟等[7]结合景观连接度分析建立了繁殖期丹顶鹤Grus japonensis的生境适宜度指数模型,得到丹顶鹤生境景观连接度可视图,对恢复丹顶鹤生境有重大意义。冯波等[8]利用分位数回归方法建立了西南太平洋阿根廷滑柔鱼Illex argentinus的HSI模型,利用Marine Explorer 4.0软件绘制了阿根廷滑柔鱼的HSI空间分布地图并加以分析。易雨君等[9]应用HSI模型及二维k-ε紊流数学模型,研究了生态因素对长江中华鲟Acipenser sinensis Gray生存、繁殖的影响。王家樵等[10]应用HSI模型和地理信息系统 (GIS)图形显示技术研究了印度洋大眼金枪鱼Thunnus obesus的栖息地分布,得出了印度洋大眼金枪鱼的最适栖息海域。对于HSI模型的研究多集中于陆地动物及鱼类,关于底栖贝类、棘皮类等海珍品的HSI模型至今尚未见报道。
本研究中,以大长山岛海域为例,根据刺参对环境的需求,建立了刺参的栖息地适宜度指数模型,对大长山岛海域的刺参生存适宜度进行了初步评价,旨在为刺参养殖区域的选择提供参考。
1 研究区域及调查站位的设置
长山群岛位于辽东半岛东南的黄海北部海域(38°13′~39°34′N,122°12′~123°13′E),该海域具有营养盐丰富、海水能见度高和生物多样性指数高等特征[11],属于典型温带季风性气候。该地区年平均气温为10℃左右,全年降水量为640 mm。长山群岛海域水深一般小于50 m,海区污染指数A值小于0.5,属无污染Ⅰ类海域,并且海域广阔,25~40 m深度范围内适宜渔业发展的海域面积可达358 828 hm2,适宜浮筏和底播养殖的海域面积为128 578 hm2[12],是中国最大的虾夷扇贝养殖基地[13]。
本研究区域位于大长山岛北部 (39°16.7′~39°17.6′N,122°37.7′~122°39.9′E),共设置7个站位,调查时间为2013年5月、8月、11月及2014年3月、5月、8月和11月 (表1、图1)。
表1 调查站位基本信息
Tab.1 The basic information on surveyed stations
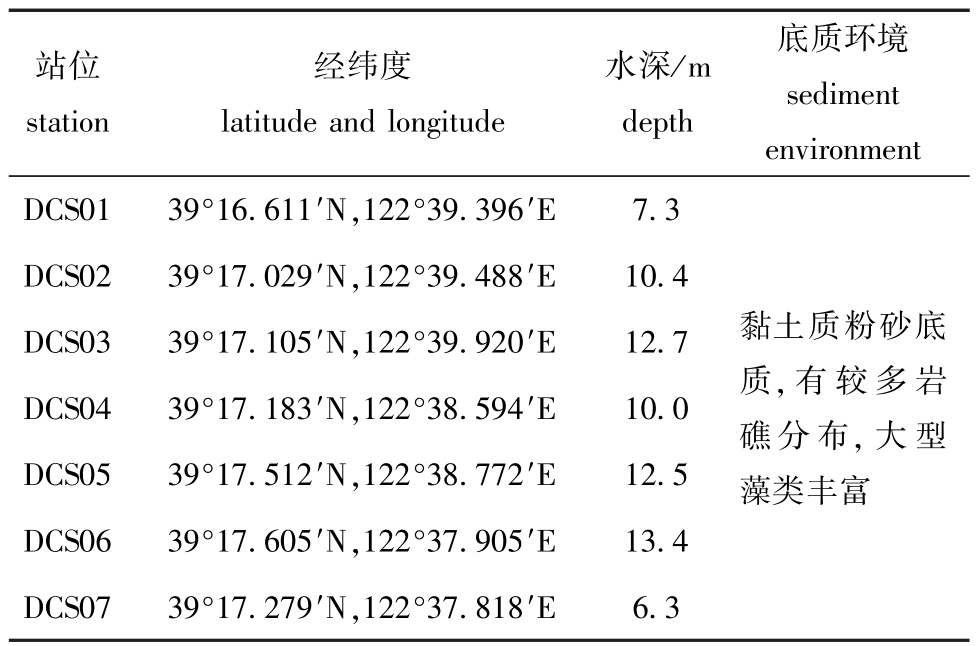
站位station经纬度latitude and longitude水深/m depth底质环境sediment environment DCS01 39°16.611′N,122°39.396′E 7.3 DCS02 39°17.029′N,122°39.488′E 10.4 DCS03 39°17.105′N,122°39.920′E 12.7 DCS04 39°17.183′N,122°38.594′E 10.0 DCS05 39°17.512′N,122°38.772′E 12.5 DCS06 39°17.605′N,122°37.905′E 13.4 DCS07 39°17.279′N,122°37.818′E 6.3黏土质粉砂底质,有较多岩礁分布,大型藻类丰富
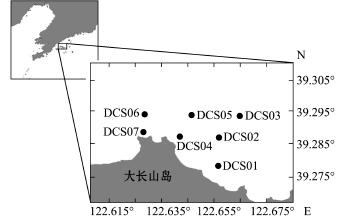
图1 研究区域及调查站位分布图
Fig.1 Area surveyed and distribution of sampling stations
2 刺参栖息地适宜度指数模型
采用赋予权重的几何平均法进行计算栖息地指数的计算,公式如下:
其中:SIi为取值为0~1的第i个影响因子的适宜度指数,0表示不适宜,1表示非常适宜;wi为第i个影响因子对应的权重值,权重越大,表明该因子对刺参的影响程度越高。
刺参的生长繁殖受众多因素的影响,主要与水温、盐度、溶解氧含量和底质类型有关。本研究中选取了水温、盐度、溶解氧、底质环境、底质中砾石与砂含量5个主要影响因子 (表2),采用层次分析法计算各因子的权重。通过对大量资料进行分析,确定各影响因子的单因素适宜度指数SI,并绘制适宜度指数曲线图。通过分析,得出影响刺参栖息环境的影响因子适宜度曲线如图2所示。
3 结果
运用HSI计算公式对大长山海域2013—2014年度共7个航次的调查影响因子实测数据进行适宜度指数计算,得出了刺参底栖环境HSI值如表3所示。利用SPSS 19软件对计算结果进行方差分析,结果表明,7个航次中,2013年8月与2014年3月的HSI平均值分别与其他6个航次有极显著性差异 (P<0.01),2013年5月、11月及2014年5月、8月和11月这5个航次之间无显著性差异(P>0.05)(表3)。
根据2013—2014年大长山研究海域刺参适宜度指数平均值变化来看:5月 (春季)及11月(秋季)适宜度指数相对较高,其中两年5月的适宜度指数均为0.66,11月分别为0.67和0.65;8 月 (夏季)次之,两年的平均值分别为0.59和0.65;3月 (冬季)最低,为0.46(表3)。
表2 影响刺参栖息地适宜度的单因素研究结果
Tab.2 The review of single factor impacting the habitat suitability of sea cucumber

影响因子impact factor权重weight说明explanation水温 0.1747陈勇等[14]研究认为,刺参的适宜水温为9~21℃,15℃时刺参体质量增长最快。董云伟等[15]研究表明,温度对刺参幼参生长有显著影响,最适生长温度为15.5℃。王国利等[16]研究表明,刺参特定生长率在17℃时达到最大值,温度超过23.6℃时,刺参出现负增长盐度 0.0655王国利等[16]研究表明,盐度对刺参的生长有明显的影响,刺参生长率在盐度为23时最低,在盐度为29时生长率最高。袁秀堂[17]研究推断,刺参的生长最适盐度范围为29~30,盐度为22.0~31.5范围内,刺参饲料转化效率和表观消化率随盐度的升高而增大溶解氧含量 0.0903刺参耐低氧能力较强[1],在正常环境中不易受到缺氧的危害。大多数底栖动物致死溶解氧浓度为2.15 mg/L[18]。刘国山等[19]在调查威海人工鱼礁区刺参大面积死亡原因中发现,底层海水缺氧可能是导致刺参死亡的主要原因底质环境 0.4196刺参多生活在海流顺畅、饵料丰富,岩礁、乱石和泥沙相间的底质或藻类繁生的泥沙底质中[1,20]。李元山等[21]调查刺参的生活环境时发现,在海藻繁茂、敌害生物少的岩礁底质上刺参分布密度较大,而粒径较小的软泥底质不适合刺参栖息底质中砾石、砂含量 0.2499泥沙混合、细砂与粉砂含量少的底质,海参分布密度大,含泥量超过20%以上的多泥底质中很少有海参栖息[20]。赵鹏等[22]通过比较刺参肠道内含物及自然沉积物的粒度来研究沉积物粒度与刺参分布之间的关系,结果表明,相比于以粉砂、细沙和中砂为主的海底,刺参倾向于分布在粗砂、极粗砂和砾石比例较高的区域。周玮等[23]对自然海域及池塘养殖的仿刺参消化道内含物中砂砾粒度进行调查并做了相关试验,结果表明,仿刺参对摄食砂砾的选择粒度集中在0.25~0.50 mm;而自然海区及池塘养殖的仿刺参,对粒度为0.125~0.25 mm、0.25~0.50 mm的选择性均高于其他粒度
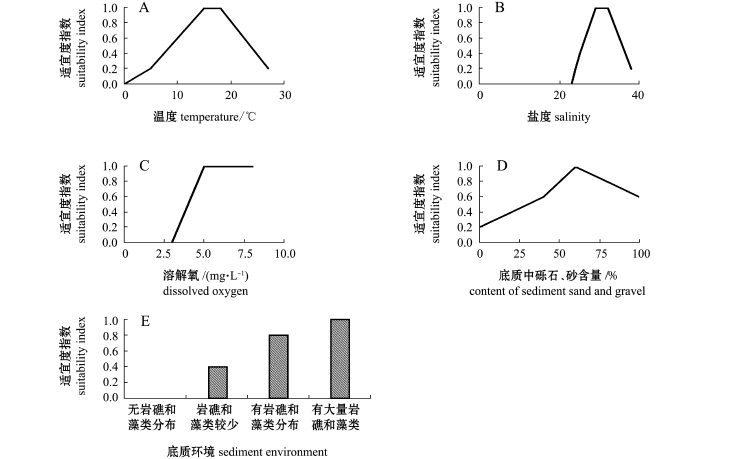
图2 刺参各影响因子的适宜度指数曲线
Fig.2 Suitability index curves of habitat for sea cucumber
各航次中,DCS01、DCS02和DCS07站位的HSI值较高。以2014年11月航次为例,DCS01、DCS02和DCS07站位的适宜度指数分别为0.66、0.67和0.67,其他4个站位的适宜度指数分别为0.65、0.62、0.63和0.62。
表3 HSI值计算结果
Tab.3 Values of habitat suitability index
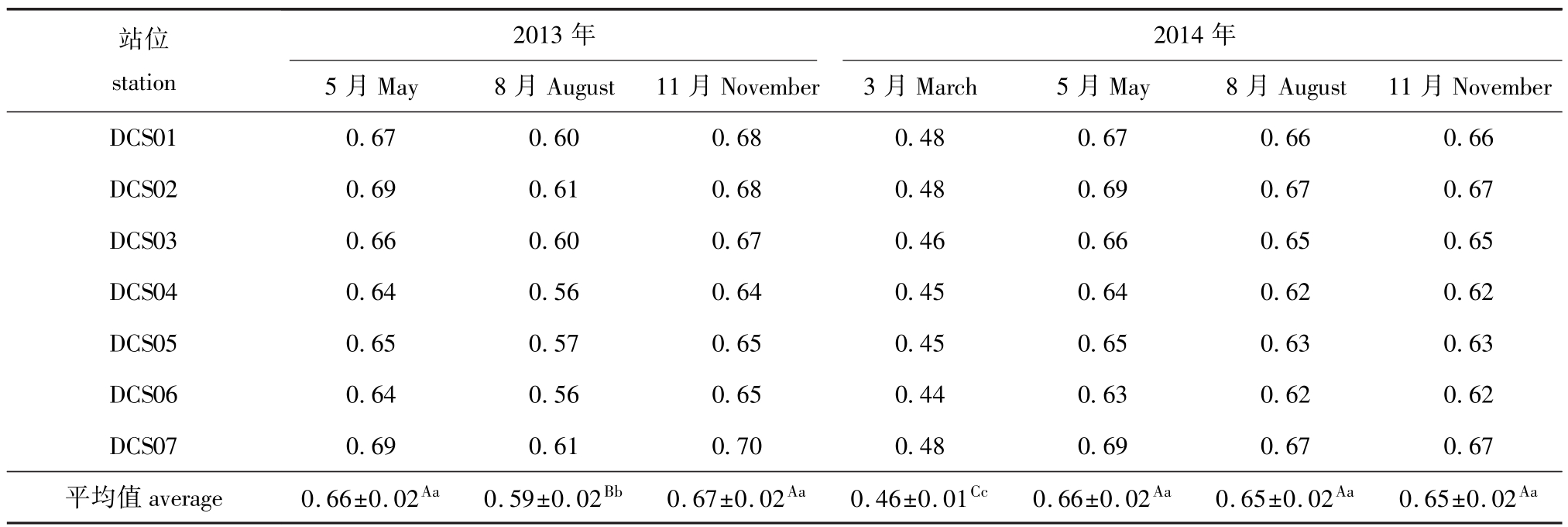
注:同行中标有不同大写字母者表示组间有极显著性差异 (P<0.01),标有不同小写字母者表示组间有显著性差异 (P<0.05),标有相同小写字母者表示组间无显著性差异 (P>0.05)
Note:The means with different capital letters within the same line are very significantly different at the 0.01 probability level,with different letters being significantly different at the 0.05 probability level,and the means with the same letters within the same line are not significant differences
站位station 2013年2014年5月May 8月August 11月November 3月March 5月May 8月August 11月November DCS01 0.67 0.60 0.68 0.48 0.67 0.66 0.66 DCS02 0.69 0.61 0.68 0.48 0.69 0.67 0.67 DCS03 0.66 0.60 0.67 0.46 0.66 0.65 0.65 DCS04 0.64 0.56 0.64 0.45 0.64 0.62 0.62 DCS05 0.65 0.57 0.65 0.45 0.65 0.63 0.63 DCS06 0.64 0.56 0.65 0.44 0.63 0.62 0.62 DCS07 0.69 0.61 0.70 0.48 0.69 0.67 0.67平均值average 0.66±0.02Aa 0.59±0.02Bb 0.67±0.02Aa 0.46±0.01Cc 0.66±0.02Aa 0.65±0.02Aa 0.65±0.02Aa
4 讨论
大量研究表明,温度是影响刺参生长的限制因子。于金海等[24]认为,仿刺参在-1.2~30.4℃温度范围内可以存活;于东祥等[25]研究表明,在小于5℃的水温环境下,幼参摄食减少,生长缓慢。本研究中,从全年HSI值的变化可以推断,大长山海域春季及秋季适宜度指数相对较高,夏季次之,冬季最低。在其他因素相同的情况下,春、秋季水温为11~13℃,适宜刺参生长,故适宜度指数较高;冬季水温为2~3℃,刺参生长减缓,故适宜度指数较低。刺参在夏季水温升高到一定范围后,会迁移到海水较深、较安静的岩石间不动不食,这种现象称为 “夏眠”[26],因此,夏季的适宜度指数也因水温较高的影响而呈现出较低的数值。根据适宜度指数模型推测出的环境适宜性与相关的研究结果基本一致,证明利用模型来估测刺参对环境的适宜性是可行的。
早川豊[27]对日本青森县野边地町及川内町海参分布区的生态调查结果显示,底质中泥含量10%以下或以砂砾底为主的海域海参分布量较多。在青岛港东沿海人工苗种放流海区的调查结果也表明,海参在泥沙混合及细砂、粉砂含量少的底质中分布密度较大[20]。本研究中,HSI的计算结果表明,DCS01、DCS02和DCS07站位的适宜度指数较高。根据大长山海域的环境调查数据,各调查站位均有大量礁石、藻类分布,且温度、盐度等其他指标无明显差异,但DCS01、DCS02和DCS07站位底质的砂砾含量要高于其他站位,这类底质更适宜刺参生活。可以推断,底质砂砾含量的差异致使这3个站位的适宜度较高,在这3个站位进行刺参苗种底播,苗种的成活率会更高,长势也会更好。
5 结语
本研究结果表明,利用栖息地适宜度指数模型这种生境评估方法,可以对刺参底播时间和地点进行更合理地规划,也可以根据不同海域的具体情况来调整模型,使这种评价方法广泛应用到不同地区。但目前关于刺参在自然环境中生活的基础生物学研究尚不全面,环境是否适宜刺参生活的判断标准缺乏确凿的科学依据作为支撑,大部分只能依靠经验赋值、主观判断来确定。因而,后续应加强对刺参等底播经济种类底栖生态学方面的基础研究,深入考虑在模型中加入海水流速、光照强度等评价指标,并利用潜水摄像、潜水采样等技术来验证模型,使优化后的模型判定结果更具有生态学价值和实践意义。
参考文献:
[1] 常亚青,丁君,宋坚,等.海参、海胆生物学研究与养殖[M].北京:海洋出版社,2004.
[2] 金龙如,孙克萍,贺红士,等.生境适宜度指数模型研究进展[J].生态学杂志,2008,27(5):841-846.
[3] 龚彩霞.基于栖息地指数的西北太平洋柔鱼渔获量估算[D].上海:上海海洋大学,2012.
[4] Vinagre C,Fonseca V,Cabral H,et al.Habitat suitability index models for the juvenile soles,Solea solea and Solea senegalensis,in the Tagus estuary:defining variables for species management[J]. Fisheries Research,2006,82(1-2):140-149.
[5] Gore J A,Bryant Jr R M.Temporal shifts in physical habitat of the crayfish,Orconectes neglectus(Faxon)[J].Hydrobiologia,1990,199(2):131-142.
[6] Soniat T M,Brody M S.Field validation of a habitat suitability index model for the American oyster[J].Estuaries,1988,11(2):87-95.
[7] 朱丽娟,刘红玉.挠力河流域丹顶鹤繁殖期生境景观连接度分析[J].生态与农村环境学报,2008,24(2):12-16,83.
[8] 冯波,田思泉,陈新军.基于分位数回归的西南太平洋阿根廷滑柔鱼栖息地模型研究[J].海洋湖沼通报,2010(1):15-22.
[9] 易雨君,王兆印,姚仕明.栖息地适合度模型在中华鲟产卵场适合度中的应用[J].清华大学学报:自然科学版,2008,48 (3):340-343.
[10] 王家樵,朱国平,许柳雄.基于HSI模型的印度洋大眼金枪鱼栖息地研究[J].海洋环境科学,2009,28(6):739-742.
[11] 王恒.国家海洋公园选址研究——以大连长山群岛为例[J].自然资源学报,2013,28(3):492-503.
[12] 张耀光,孙才志,刘锴,等.海岛海域生物资源利用与海洋农牧化生产布局新发展的研究——以长山群岛为例[J].自然资源学报,2009,24(6):945-955.
[13] 马玉艳,韩华,卞正和,等.大小长山岛周围海域浮游动物群落分布特征[J].海洋环境科学,2012,31(3):364-369.
[14] 陈勇,高峰,刘国山,等.温度、盐度和光照周期对刺参生长及行为的影响[J].水产学报,2007,31(5):687-691.
[15] 董云伟,董双林,田相利,等.不同水温对刺参幼参生长、呼吸及体组成的影响[J].中国水产科学,2005,12(1):33-37.
[16] 王国利,祝文兴,李兆智,等.温度与盐度对刺参(Apostichopus japonicus)生长的影响[J].山东科学,2007,20(3):6-9.
[17] 袁秀堂.刺参Apostichopus japonicus(Selenka)生理生态学及其生物修复作用的研究[D].青岛:中国科学院海洋研究所,2005.
[18] 丸茂恵右,横田瑞郎.貧酸素水塊の形成および貧酸素の生物影響た関する文献調査[J].海生研研報,2012(15):1-21.
[19] 刘国山,蔡星媛,佟飞,等.威海双岛湾人工鱼礁区刺参大面积死亡原因初探[J].渔业信息与战略,2014,29(2):122-129.
[20] 于东祥,孙慧玲,陈四清,等.海参健康养殖技术[M].2版.北京:海洋出版社,2010.
[21] 李元山,王远隆,王培良.刺参的生物环境及增殖水域选择[J].海洋湖沼通报,1994(4):42-47.
[22] 赵鹏,杨红生.刺参对不同生境中沉积物粒度的选择[J].海洋科学,2010,34(4):11-16.
[23] 周玮,王俊杰,陆佳,等.仿刺参摄食砂砾粒度的选择性研究[J].大连水产学院学报,2009,23(6):446-450.
[24] 于金海,冷晓飞,李春艳,等.黄海北部仿刺参养殖池塘关键环境因子的周年变化与管理研究[J].大连水产学院学报,2009,24(6):514-519.
[25] 于东祥,宋本祥.池塘养殖刺参幼参的成活率变化和生长特点[J].中国水产科学,1999,6(3):109-110.
[26] 袁秀堂,杨红生,陈慕雁,等.刺参夏眠的研究进展[J].海洋科学,2007,31(8):88-90.
[27] 早川豊.マナマコ生態調査[J].青森県水産増殖センター事業概要,1978,7:173-184.
基于Ecopath模型的獐子岛人工鱼礁海域生态系统结构和功能变化
许祯行,陈勇,田涛,刘永虎,尹增强,刘汉超
(大连海洋大学辽宁省海洋牧场工程技术研究中心,辽宁大连116023)
摘要:为了研究獐子岛海洋牧场的生态系统结构和功能,基于Ecopath with Ecosim 6.4软件,构建了獐子岛人工鱼礁区生态系统2010—2012年夏季的Ecopath模型,分析了3个时期海域生态系统的能量流动结构和功能变化,模型由23个功能组构成,基本覆盖了该海域生态系统能量流动的全过程。结果表明:2010—2012年夏季獐子岛海域人工鱼礁区生态系统中,虾夷扇贝Patinopecten yessoensis、许氏平鲉Sebastes schlegelii和大泷六线鱼Hexagrammos otakii等生物营养级升高;皱纹盘鲍Haliotis discus hannai的猎物重叠度指数平均值从0.123升至0.139,捕食重叠度指数平均值从0.112升至0.165,许氏平鲉的捕食重叠度指数平均值从0.063升至0.185,大泷六线鱼的捕食重叠度平均值从0.062升至0.132;该生态系统能量转化效率由2010年夏季的11.1%升至2012年夏季的14.4%;3个时期獐子岛人工鱼礁区生态系统的循环指数 (FCI)值均大于0.1,再循环率较高,系统净生产力 (NSP)值较高,杂食指数 (SOI)从0.136升至0.172,联结指数 (CI)从0.203升至0.224,平均能流路径 (MPL)从2.555升至3.486。研究表明,獐子岛人工鱼礁区海域生态系统处于尚未稳定的成熟期。
关键词:人工鱼礁;生态系统结构;能量流动;生态位重叠度;Ecopath模型
DOI:10.16535/j.cnki.dlhyxb.2016.01.015
文章编号:2095-1388(2016)01-0085-10
獐子岛海洋牧场 (东经124°47′、北纬39°3′)地处黄海北部外长山列岛的最南端,水质条件优越,营养盐丰富[1],盛产刺参、皱纹盘鲍和虾夷扇贝等多种海产品,是中国规模最大的海洋牧场示范基地。近些年,獐子岛海洋牧场进行了大规模的人工鱼礁建设,以改善和优化生态环境,养护和增殖渔业资源。目前,对獐子岛海洋牧场的研究大多集中在人工鱼礁区增养殖、生态环境与水质环境的变化等方面。张继红等[2]研究了獐子岛海域虾夷扇贝的养殖容量,认为在虾夷扇贝养殖中32亿粒接近该海域的生态容量。于莹等[3]对獐子岛海域浮游纤毛虫丰度和生物量的研究表明,该海域2010年7月砂壳纤毛虫有17种,8月份有21种。尹洁慧[4]对獐子岛海域浮游动物群落时空变化特征及其与环境因子之间关系的研究表明,淡水的输入与积累营养盐的黄海冷水团可促进浮游植物的初级生产。王全超等[5]对獐子岛马牙滩潮间带及近岸海区大型底栖动物群落特征的研究表明,大型底栖动物群落结构处于不稳定状态。陈勇等[6]对獐子岛海洋牧场人工鱼礁区鱼类资源养护效果的研究表明,人工鱼礁区鱼类与投礁前相比增殖效果明显,资源养护效果较好。但对该海域生态系统结构和功能量化的系统研究目前尚未见报道。
Ecopath模型可以量化生态系统中众多指标,被国际专家学者认为是未来水域生态系统研究的核心工具[7],有学者也已利用该模型对人工鱼礁区生态系统展开了类似研究。李永刚等[8]、赵静等[9]和吴忠鑫等[10]基于该模型分别对嵊泗人工鱼礁区、枸杞海藻场生态系统能量和荣成俚岛人工鱼礁区生态系统结构和功能进行了评价。本研究中,根据2010—2012年獐子岛夏季海洋资源调查数据,利用Ecopath with Ecosim(EwE)6.4软件构建了獐子岛人工鱼礁海域生态系统能量流动模型,并对该海域生态系统特征进行量化分析,探讨了獐子岛海洋牧场生态系统结构和功能的变化以及人工鱼礁建设效果,旨在为獐子岛海洋牧场的建设和管理提供科学依据,为中国近海海洋牧场建设提供参考。
收稿日期:2015-05-23
基金项目:国家 “十二五”科技支撑计划项目 (2012BAD18B03,2013BAD23B02);国家海洋公益性行业科研专项 (201205023)
作者简介:许祯行 (1990—),男,硕士研究生。E-mail:334877031@qq.com
通信作者:田涛 (1979—),男,博士,副教授。E-mail:tian2007@dlou.edu.cn
1 材料与方法
1.1 数据来源
本研究中数据源于獐子岛海域内2010年夏季本底调查以及2011年夏季与2012年夏季獐子岛人工鱼礁区资源环境养护情况跟踪调查 (图1),包括小型拖网调查、水下摄像与潜水样方调查,并对样品进行了处理和分析 (采样和分析按 《海洋调查规范》执行),满足构建Ecopath模型的基本数据需求。
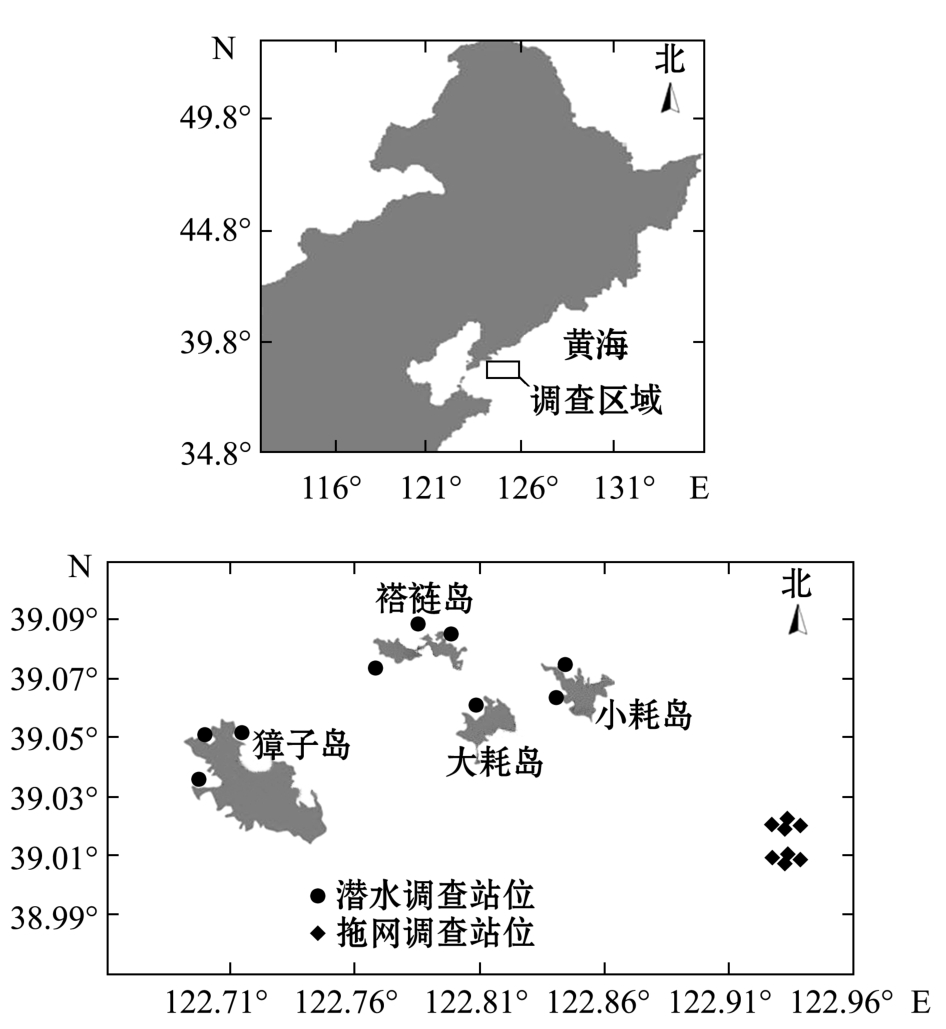
图1 调查区域及站位分布图
Fig.1 Area surveyed and distribution of sampling stations
1.2 Ecopath模型的建立
1.2.1 功能组的划分 Ecopath模型所定义的生态系统由一系列生态关联的功能组 (group)组成。为了探讨獐子岛人工鱼礁海域增殖对象生物的营养级、结构和功能的变化,本研究中将虾夷扇贝Patinopecten yessoensis、刺参Apostichopus japonicus、皱纹盘鲍 Haliotis discus hannai、许氏平鲉 Sebastes schlegelii、大泷六线鱼Hexagrammos otakii和海胆6种研究对象各自划分为独立的功能组,其他生物按照规格、生物学和食性特征划分成17个功能组,共计23个功能组 (表1),这些功能组基本覆盖生态系统能量流动的全过程。
1.2.2 功能组的参数来源 功能组内生物量数据由现场调查评估所得,礁区附近的生物量采用阿氏拖网进行扫海面积法估算,网口长度为2 m,拖网时长约20 min,拖网船功率为110.3 kW,拖速约为4 kn。礁区内便于采捕的生物,如底栖藻类、刺参和皱纹盘鲍等,其生物量采用潜水样方调查法估算,其他生物的生物量采用水下摄像方法估算,浮游植物生物量通过调查所得的叶绿素a含量估算[11],有机碎屑生物量采用Pauly等[12]提出的有关初级生产碳与有机碎屑的经验公式估算。食物分析矩阵中的食性关系根据采样鱼类的胃含物分析获得,模型中生产量与生物量的比值 (P/B)、消耗量与生物量的比值 (Q/B)、异养细菌的生物量与其他动物的食性,参考附近海域和世界同纬度相近海域的Ecopath模型中类似功能组的值得出[8,13-14]。
1.2.3 生态位重叠度指数 Clarles Elton(1927)
从营养角度将生态位定义为该动物在生物环境中的地位及其食物与天敌的关系[15]。生态位重叠度指数 (niche overlap index)是指两个物种在相同资源位上相遇的频率[16]。EwE软件中使用了Pianka[17]提出的基于Lotka-Voerma方程求得的猎物重叠度指数 (prey overlap index)与捕食重叠度指数(predator overlap index),实现了功能组内生物之间食物的竞争程度与天敌重叠情况的定量分析。
猎物重叠度指数方程为
其中:Ojk为j和k物种捕食其他物种的重叠度指数;Pji和Pki分别为j和k物种捕食i物种的比例,若Pji和Pki中有估值为0的情况,则引进等差中项的概念估算Ojk, 即
捕食重叠度指数方程为
其中:Pmn为m和n物种的捕食重叠度指数;Xml和Xnl分别为m和n物种被所有捕食者l消耗的比例。
1.2.4 模型质量分析 参数来源的可靠性和准确度是影响Ecopath模型质量的主要因素。本研究中主要用Pedigree指数 (Pedigree index)量化输入模型中参数的质量和不确定性,该指数通过EwE软件中Pedigree程序提供的指数定义表量化模型中每个参数来源,然后通过公式计算得出Pedigree指数。Pedigree指数计算公式为
表1 獐子岛人工鱼礁区生态系统Ecopath模型的功能组及其主要种类组成
Tab.1 Functional groups and main species check list in artificial reef ecosystem in Zhangzi Island based on Ecopath model

编号No. 功能组functional group组成composition 1 底栖藻类和海草bethic algae and seaweed 海带 Laminaria japonica、裙带菜 Undaria pinnatifida Suringar、江蓠 Gracilaria、大叶藻Zostera marina 2 浮游植物phytoplankton 甲藻门Pyrrophyta、硅藻门Bacillariophyta、蓝藻门Cyanophyta 3 浮游动物zooplankton 桡足类copepods、甲壳类crustacea、多毛类polychetes、枝角类cladocera 4 虾夷扇贝Patinopecten yessoensis 虾夷扇贝Patinopecten yessoensis 5 皱纹盘鲍Haliotis discus hannai 皱纹盘鲍Haliotis discus hannai 6 刺参Apostichopus japonicus 刺参Apostichopus japonicus 7 许氏平鲉Sebastes schlegelii 许氏平鲉Sebastes schlegelii 8 大泷六线鱼Hexagrammos otakii 大泷六线鱼Hexagrammos otakii 9 软体动物mollusca 双壳类lamellibranchia、腹足类gastropoda 10 头足类cephalopod 短蛸Octopus ocellatus、长蛸Octopus variabilis 11 虾类shrimps 中国毛虾Acetes chinensis、脊腹褐虾Crangon affinis 12 蟹类crabs 日本蟳Charybdis japonica、枯瘦突眼蟹Oregonia gracilis、寄居蟹Pagurus sp. 13 大中型底层鱼类large bottom fishes 黄鮟鱇Lophius litulon 14 棘皮类echinodermata 罗氏海盘车 Asierias rollestoni Bell、萨氏真蛇尾Ophiura sarsii、马氏刺蛇尾Ophrothrix marenzelleri、紫蛇尾Ophiopholis mirabilis 15 其他底栖动物other benthos 多毛类polychetes、环节类annelida 16 小型底层鱼类small demersal fishes 矛尾鰕虎鱼 Chaeturichthys stigmatias Richardson、高眼鲽Cleisthenes herzensteini、方氏云鳚Enedrias fangi Wang et Wang 17 大型中上层鱼类large pelagic fishes 太平洋鲱Clupea pallasii 18 小型中上层鱼类small pelagic fishes 沙氏下鱵鱼Hyporhamphus sojori、日本鯷Engraulis japonicus 19 海胆sea urchin 光棘球海胆Strongylocentrotus nudus、虾夷马粪海胆Strongylocentrotus intermedius 20 海洋哺乳动物marine mammals 江豚Neophocaena phocaenoides 21 海鸟fish-eating birds 海鸥Larus canus 22 异养细菌heterotrophic bacteria 异养细菌heterotrophic bacteria 23 有机碎屑detritus 有机碎屑detritus
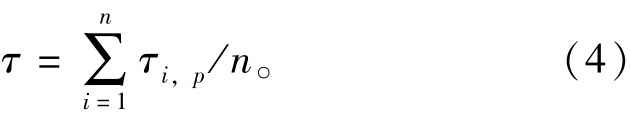
其中:τi,p为第p类i功能组中的Pedigree指数;n为功能组的总数;p为B、P/B、Q/B、食物矩阵和渔获量等输入参数的种类。经计算,本研究中2010、2011、2012年Ecopath模型的Pedigree指数分别为0.554、0.619和0.567。
1.2.5 模型的调试和输出 生态营养转化效率(EE)通过Ecopath模型估算得出,当某些功能组EE值大于1时,说明该生态系统的输入量和输出量不均等,需通过对食物分析矩阵等参数进行微调,使功能组内所有EE值均调整到小于1且P/Q<0.3,以保证生态系统能量流动的平衡。该模型经调试后得到输出结果如表2所示。
2 结果与分析
2.1 各功能组的营养级与生态营养转化效率
经计算,得出獐子岛海域生态系统各功能组估算参数如表3所示。从表3可见:2010—2012年夏季,虾夷扇贝的营养级从2.120升至2.300,许氏平鲉从3.676升至3.790,大泷六线鱼从3.210升至3.421,刺参从 2.490降至 2.408,海胆从2.047降至2.032;刺参的生态营养转化效率从0.143升至0.682,海胆从0.108升至0.601,许氏平鲉从0.107降至0.036,大泷六线鱼从0.062降至0.040。
表2 2010—2012年夏季獐子岛人工鱼礁区Ecopath模型功能组输入参数
Tab.2 Input parameters of Ecopath model in Zhangzi Island artificial reef from summer of 2010 to summer of 2012
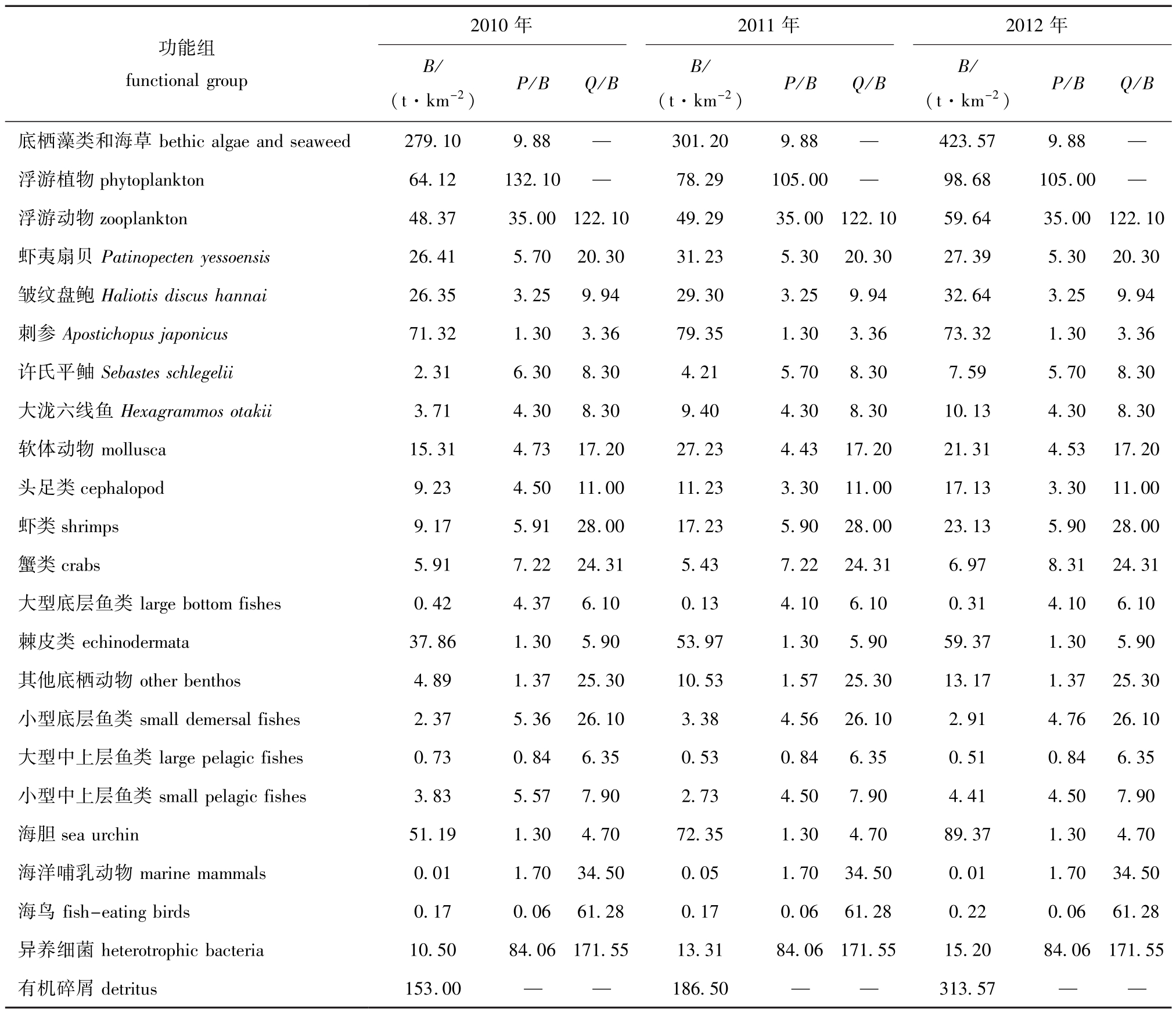
注:B为生物量;P为生产量;Q为消耗量
Note:B,biomass;P,production;Q,consumption
功能组functional group 2010年2011年2012年B/ (t·km-2)P/B Q/B B/ (t·km-2)P/B Q/B B/ (t·km-2)P/B Q/B底栖藻类和海草bethic algae and seaweed 279.10 9.88 — 301.20 9.88 — 423.57 9.88—浮游植物phytoplankton 64.12 132.10 — 78.29 105.00 — 98.68 105.00 —浮游动物zooplankton 48.37 35.00 122.10 49.29 35.00 122.10 59.64 35.00 122.10虾夷扇贝Patinopecten yessoensis 26.41 5.70 20.30 31.23 5.30 20.30 27.39 5.30 20.30皱纹盘鲍Haliotis discus hannai 26.35 3.25 9.94 29.30 3.25 9.94 32.64 3.25 9.94刺参Apostichopus japonicus 71.32 1.30 3.36 79.35 1.30 3.36 73.32 1.30 3.36许氏平鲉Sebastes schlegelii 2.31 6.30 8.30 4.21 5.70 8.30 7.59 5.70 8.30大泷六线鱼Hexagrammos otakii 3.71 4.30 8.30 9.40 4.30 8.30 10.13 4.30 8.30软体动物mollusca 15.31 4.73 17.20 27.23 4.43 17.20 21.31 4.53 17.20头足类cephalopod 9.23 4.50 11.00 11.23 3.30 11.00 17.13 3.30 11.00虾类shrimps 9.17 5.91 28.00 17.23 5.90 28.00 23.13 5.90 28.00蟹类crabs 5.91 7.22 24.31 5.43 7.22 24.31 6.97 8.31 24.31大型底层鱼类large bottom fishes 0.42 4.37 6.10 0.13 4.10 6.10 0.31 4.10 6.10棘皮类echinodermata 37.86 1.30 5.90 53.97 1.30 5.90 59.37 1.30 5.90其他底栖动物other benthos 4.89 1.37 25.30 10.53 1.57 25.30 13.17 1.37 25.30小型底层鱼类small demersal fishes 2.37 5.36 26.10 3.38 4.56 26.10 2.91 4.76 26.10大型中上层鱼类large pelagic fishes 0.73 0.84 6.35 0.53 0.84 6.35 0.51 0.84 6.35小型中上层鱼类small pelagic fishes 3.83 5.57 7.90 2.73 4.50 7.90 4.41 4.50 7.90海胆sea urchin 51.19 1.30 4.70 72.35 1.30 4.70 89.37 1.30 4.70海洋哺乳动物marine mammals 0.01 1.70 34.50 0.05 1.70 34.50 0.01 1.70 34.50海鸟fish-eating birds 0.17 0.06 61.28 0.17 0.06 61.28 0.22 0.06 61.28异养细菌heterotrophic bacteria 10.50 84.06 171.55 13.31 84.06 171.55 15.20 84.06 171.55有机碎屑detritus 153.00 — — 186.50 — — 313.57— —
2.2 各营养级的能流特征
本研究中,通过林德曼椎型图 (Lindeman spine),将复杂的食物网关系转换成一系列离散营养级[18],并详细标注了每个营养级的流动情况(图2),结果表明,獐子岛人工鱼礁区生态系统中同时存在碎屑食物链和牧食食物链,该海域3个时期能量流动主要在5个营养级之间发生 (图3~图5)。
3个时期,初级生产者总转化效率分别为11.1%、12.6%和14.5%,碎屑营养转化总效率分别为11.1%、12.1%和14.1% (表4),总体均呈逐渐升高的趋势。
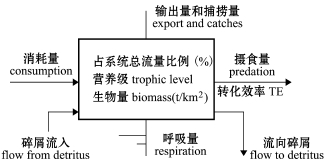
图2 林德曼椎型图中各位置的参数说明
Fig.2 Instructions of the data in Lindeman spine diagram
2.3 各功能组的生态重叠度
通过计算各功能组 (非生物与植物除外)与其他功能组生态位重叠度指数平均值的变化 (表5)发现:2010年—2012年夏季,猎物重叠度指数中,皱纹盘鲍的平均值从0.123升至0.139,虾夷扇贝从 0.310升至 0.332,虾类从 0.329升至0.410,蟹类从0.268升至0.315;捕食重叠度指数中,浮游动物的平均值从0.095升至0.163,虾夷扇贝从0.132升至0.240,皱纹盘鲍从0.112升至0.165,许氏平鲉从0.063升至0.185,大泷六线鱼从0.062升至0.132。
表3 2010—2012年夏季獐子岛人工鱼礁区Ecopath模型功能组估算参数
Tab.3 Functional parameters of Ecopath model in Zhangzi Island artificial reef from summer of 2010 to summer of 2012
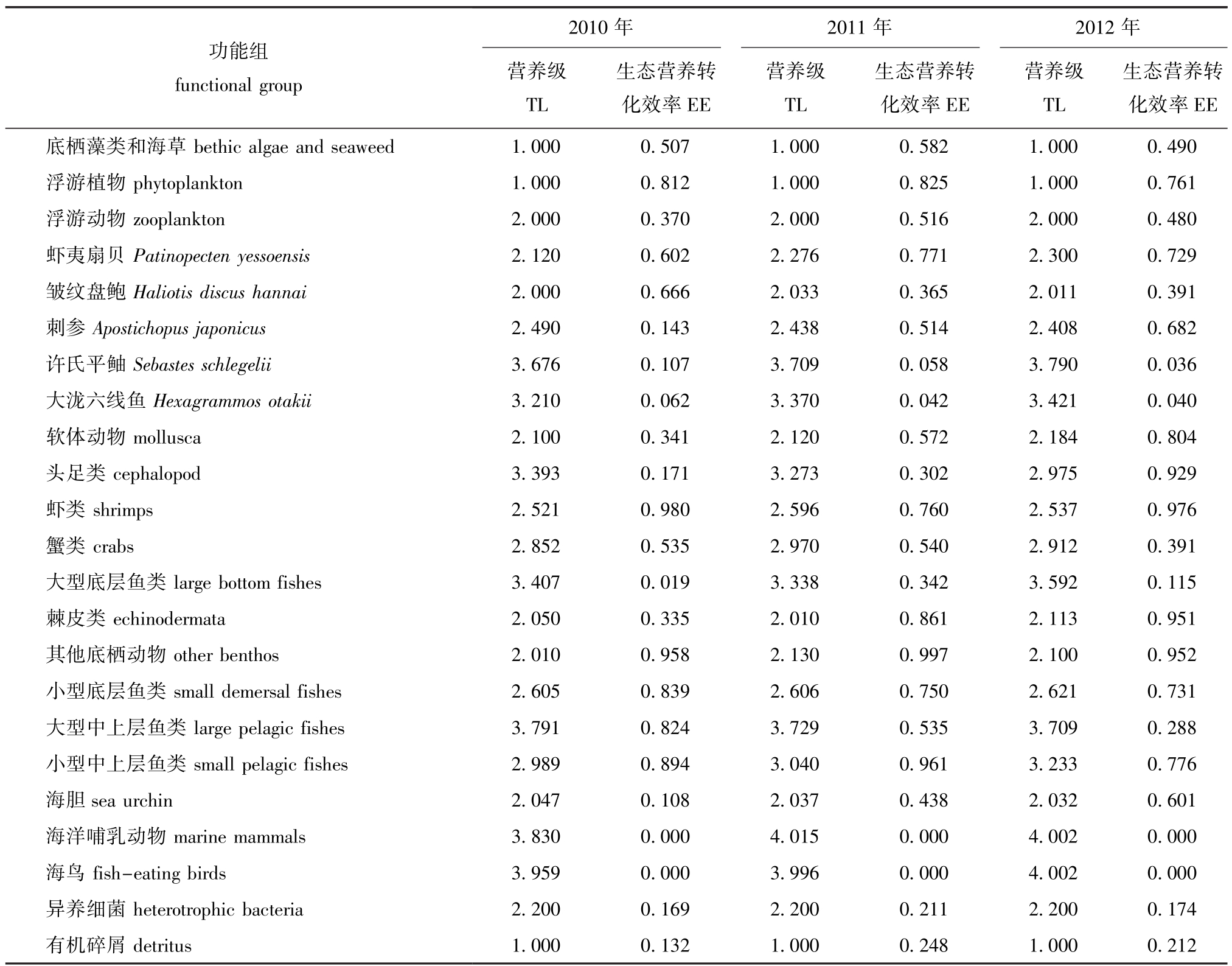
功能组functional group 2010年2011年2012年营养级TL生态营养转化效率EE营养级TL生态营养转化效率EE营养级TL生态营养转化效率EE底栖藻类和海草bethic algae and seaweed 1.000 0.507 1.000 0.582 1.000 0.490浮游植物phytoplankton 1.000 0.812 1.000 0.825 1.000 0.761浮游动物zooplankton 2.000 0.370 2.000 0.516 2.000 0.480虾夷扇贝Patinopecten yessoensis 2.120 0.602 2.276 0.771 2.300 0.729皱纹盘鲍Haliotis discus hannai 2.000 0.666 2.033 0.365 2.011 0.391刺参Apostichopus japonicus 2.490 0.143 2.438 0.514 2.408 0.682许氏平鲉Sebastes schlegelii 3.676 0.107 3.709 0.058 3.790 0.036大泷六线鱼Hexagrammos otakii 3.210 0.062 3.370 0.042 3.421 0.040软体动物mollusca 2.100 0.341 2.120 0.572 2.184 0.804头足类cephalopod 3.393 0.171 3.273 0.302 2.975 0.929虾类shrimps 2.521 0.980 2.596 0.760 2.537 0.976蟹类crabs 2.852 0.535 2.970 0.540 2.912 0.391大型底层鱼类large bottom fishes 3.407 0.019 3.338 0.342 3.592 0.115棘皮类echinodermata 2.050 0.335 2.010 0.861 2.113 0.951其他底栖动物other benthos 2.010 0.958 2.130 0.997 2.100 0.952小型底层鱼类small demersal fishes 2.605 0.839 2.606 0.750 2.621 0.731大型中上层鱼类large pelagic fishes 3.791 0.824 3.729 0.535 3.709 0.288小型中上层鱼类small pelagic fishes 2.989 0.894 3.040 0.961 3.233 0.776海胆sea urchin 2.047 0.108 2.037 0.438 2.032 0.601海洋哺乳动物marine mammals 3.830 0.000 4.015 0.000 4.002 0.000海鸟fish-eating birds 3.959 0.000 3.996 0.000 4.002 0.000异养细菌heterotrophic bacteria 2.200 0.169 2.200 0.211 2.200 0.174有机碎屑detritus 1.000 0.132 1.000 0.248 1.000 0.212
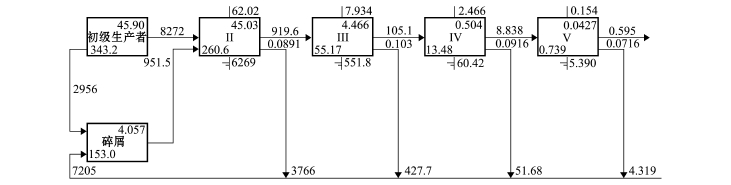
图3 2010年夏季獐子岛人工鱼礁区生态系统林德曼椎型示意图
Fig.3 Lindeman spine diagram in Zhangzi Island artificial reef ecosystem in summer of 2010
2.4 生态系统的总体特征
本研究中通过Ecopath模型估算了2010—2012年獐子岛人工鱼礁区生态系统特征参数 (表6)。系统总流量是表征系统总体规模的重要指标,从表6可见,2010—2012年系统总流量降低了33.99%。与2010年相比,2012年联结指数、系统杂食指数、循环指数和平均能流路径分别从0.203、0.136、 2.180 和 2.555 增 至 0.224、0.172、9.120和3.486,总初级生产力/总呼吸量从2.291降至2.197,3个时期系统净生产力平均值为6578.85 t/km2。
通过SPSS 13.0软件将3个时期的特征参数进行线性回归分析,结果表明,总消耗量、总呼吸量和总生物量均呈线性增长。
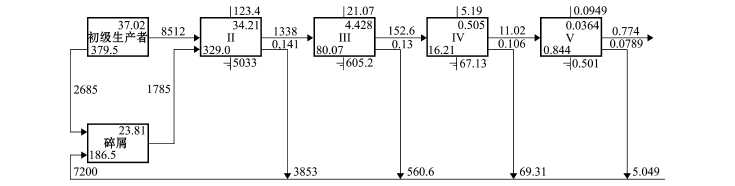
图4 2011年夏季獐子岛人工鱼礁区生态系统林德曼椎型示意图
Fig.4 Lindeman spine diagram in Zhangzi Island artificial reef ecosystem in summer of 2011
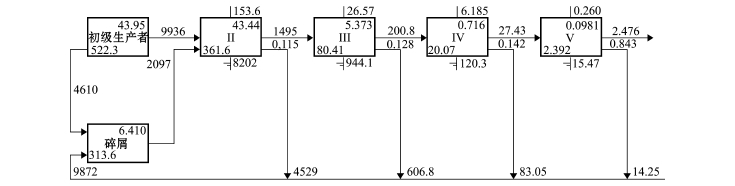
图5 2012年夏季獐子岛人工鱼礁区生态系统林德曼椎型示意图
Fig.5 Lindeman spine diagram in Zhangzi Island artificial reef ecosystem in summer of 2012
表4 2010—2012年夏季獐子岛人工鱼礁区生态系统各营养级的转化效率
Tab.4 Transfer efficiency of discrete trophic levels in Zhangzi Island artificial reef ecosystem from summer of 2010 to summer of 2012 %
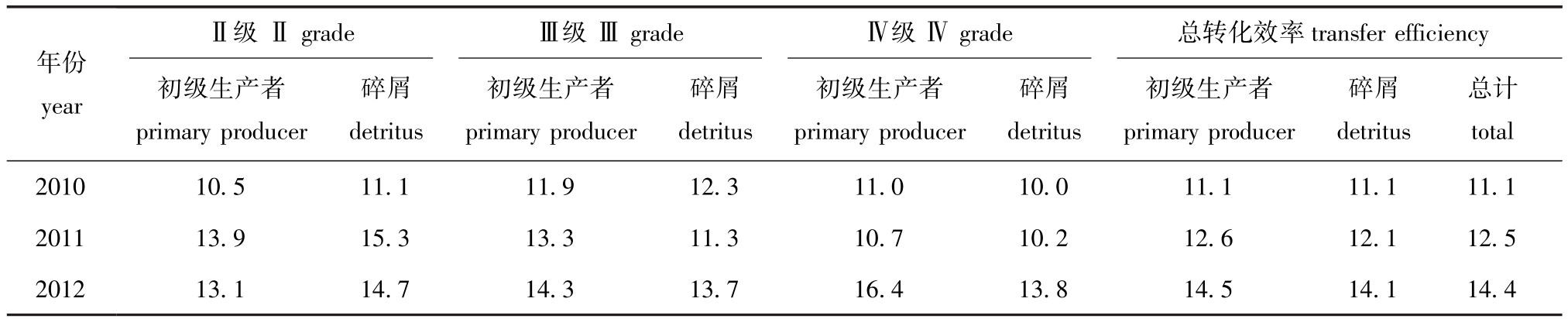
注:总转化效率是指第Ⅱ营养级到第Ⅳ营养级的几何平均数
Note:The efficiency is calculated as geometric mean from trophic levelⅡto trophic levelⅣ
年份yearⅡ级Ⅱgrade Ⅲ级Ⅲgrade Ⅳ级Ⅳgrade 总转化效率transfer efficiency初级生产者primary producer碎屑detritus初级生产者primary producer碎屑detritus初级生产者primary producer碎屑detritus初级生产者primary producer碎屑detritus总计total 2010 10.5 11.1 11.9 12.3 11.0 10.0 11.1 11.1 11.1 2011 13.9 15.3 13.3 11.3 10.7 10.2 12.6 12.1 12.5 2012 13.1 14.7 14.3 13.7 16.4 13.8 14.5 14.1 14.4
3 讨论
3.1 营养级与生态营养转化效率的变化
许氏平鲉、大泷六线鱼、皱纹盘鲍、刺参和海胆是獐子岛人工鱼礁区海域的主要增殖对象。模型运行结果表明,随着许氏平鲉和大泷六线鱼营养级水平的升高,生态营养转化效率逐渐降低 (图6、图7),说明许氏平鲉和大泷六线鱼的生态营养转化效率与营养级呈负相关关系,这与唐启升等[19]的研究中二者的变动情况基本吻合。唐启升等[19]指出,环境因素和生态生理因素是引起鱼类生理特征及营养动力学特征改变的主要因素;张波等[20]认为,同种类个体大小以及被摄食食物种类的组成变化是引起营养级波动的主要原因。人工鱼礁的投放改善了生态环境,为鱼类提供了良好的庇护场所及丰富的饵料来源,同时也为礁区内生物带来了较好的养护环境,使许氏平鲉和大泷六线鱼等生物营养级升高。
表5 2010—2012年夏季獐子岛鱼礁区生态重叠度指数平均值
Tab.5 Mean value of overlap index in Zhangzi Island artificial reef ecosystem from summer of 2010 to summer of 2012
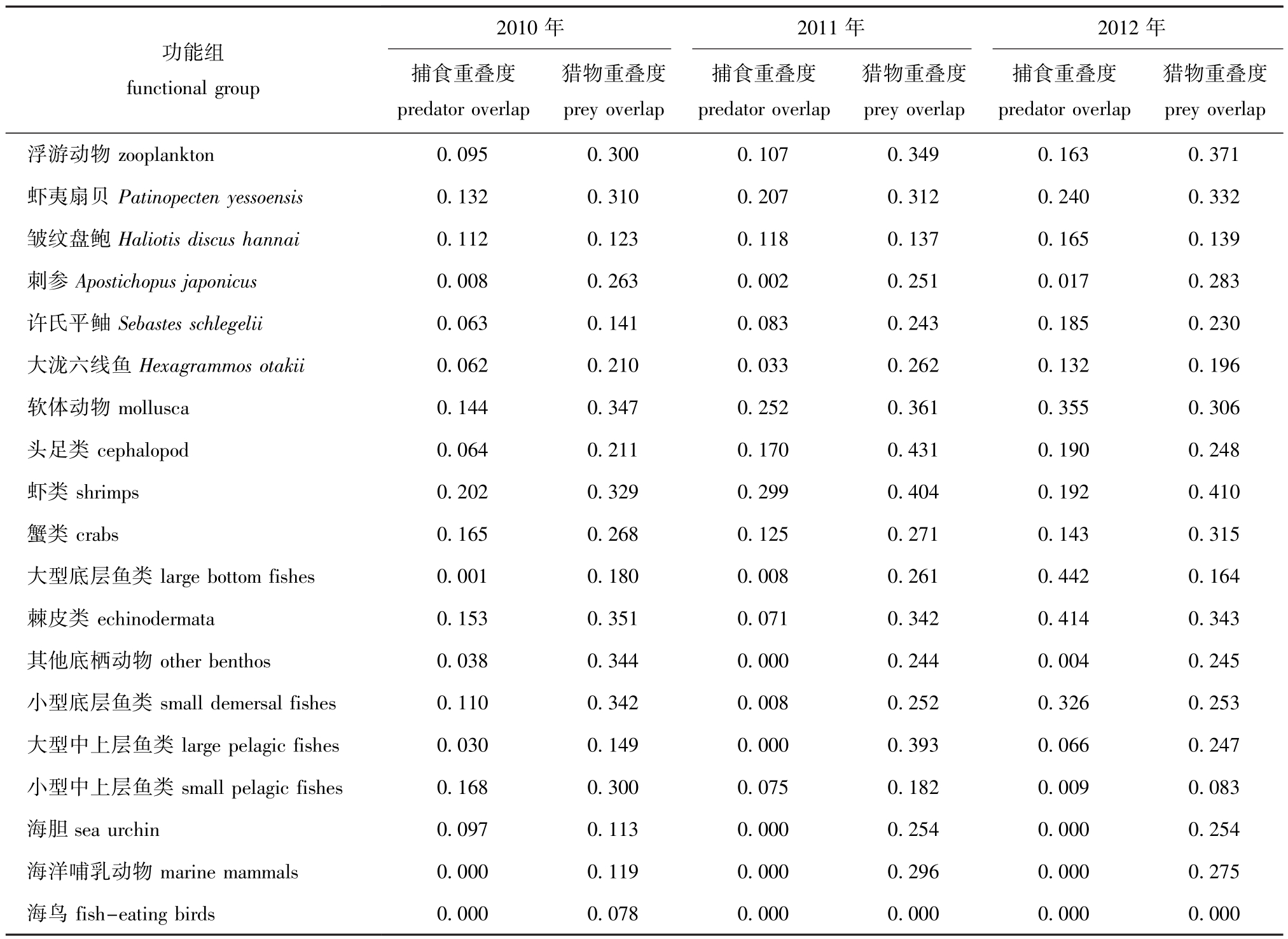
功能组functional group 2010年2011年2012年捕食重叠度predator overlap猎物重叠度prey overlap捕食重叠度predator overlap猎物重叠度prey overlap捕食重叠度predator overlap猎物重叠度prey overlap浮游动物zooplankton 0.095 0.300 0.107 0.349 0.163 0.371虾夷扇贝Patinopecten yessoensis 0.132 0.310 0.207 0.312 0.240 0.332皱纹盘鲍Haliotis discus hannai 0.112 0.123 0.118 0.137 0.165 0.139刺参Apostichopus japonicus 0.008 0.263 0.002 0.251 0.017 0.283许氏平鲉Sebastes schlegelii 0.063 0.141 0.083 0.243 0.185 0.230大泷六线鱼Hexagrammos otakii 0.062 0.210 0.033 0.262 0.132 0.196软体动物mollusca 0.144 0.347 0.252 0.361 0.355 0.306头足类cephalopod 0.064 0.211 0.170 0.431 0.190 0.248虾类shrimps 0.202 0.329 0.299 0.404 0.192 0.410蟹类crabs 0.165 0.268 0.125 0.271 0.143 0.315大型底层鱼类large bottom fishes 0.001 0.180 0.008 0.261 0.442 0.164棘皮类echinodermata 0.153 0.351 0.071 0.342 0.414 0.343其他底栖动物other benthos 0.038 0.344 0.000 0.244 0.004 0.245小型底层鱼类small demersal fishes 0.110 0.342 0.008 0.252 0.326 0.253大型中上层鱼类large pelagic fishes 0.030 0.149 0.000 0.393 0.066 0.247小型中上层鱼类small pelagic fishes 0.168 0.300 0.075 0.182 0.009 0.083海胆sea urchin 0.097 0.113 0.000 0.254 0.000 0.254海洋哺乳动物marine mammals 0.000 0.119 0.000 0.296 0.000 0.275海鸟fish-eating birds 0.000 0.078 0.000 0.000 0.000 0.000
表6 2010—2012年夏季獐子岛人工鱼礁区生态系统总体统计学比较
Tab.6 Comparison of system statistics in Zhangzi Island artificial reef ecosystem from summer of 2010 to summer of 2012
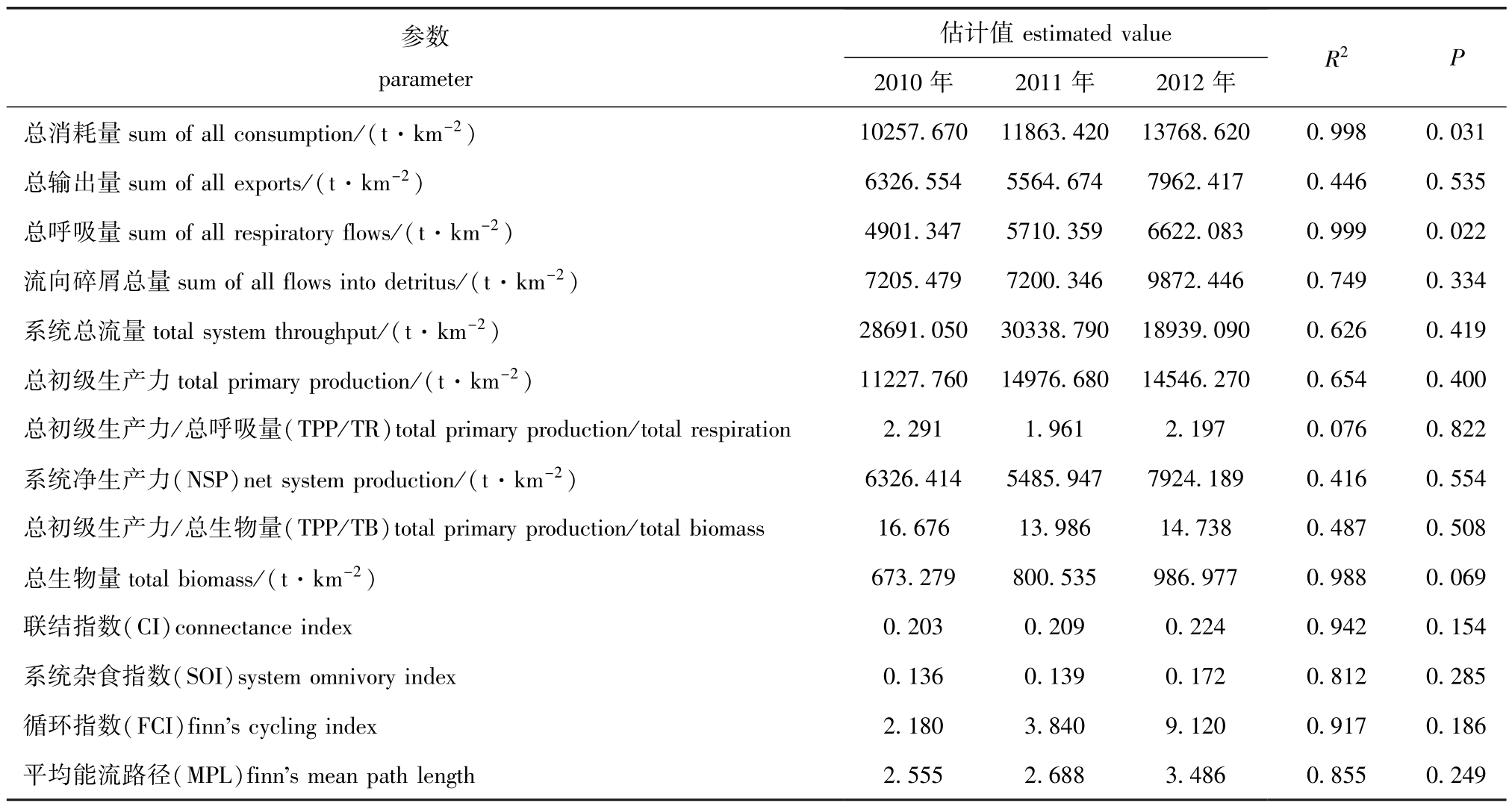
参数parameter估计值estimated value 2010年 2011年 2012年R2P总消耗量sum of all consumption/(t·km-2)10257.670 11863.420 13768.620 0.998 0.031总输出量sum of all exports/(t·km-2) 6326.554 5564.674 7962.417 0.446 0.535总呼吸量sum of all respiratory flows/(t·km-2) 4901.347 5710.359 6622.083 0.999 0.022流向碎屑总量sum of all flows into detritus/(t·km-2) 7205.479 7200.346 9872.446 0.749 0.334系统总流量total system throughput/(t·km-2) 28691.050 30338.790 18939.090 0.626 0.419总初级生产力total primary production/(t·km-2) 11227.760 14976.680 14546.270 0.654 0.400总初级生产力/总呼吸量(TPP/TR)total primary production/total respiration 2.291 1.961 2.197 0.076 0.822系统净生产力(NSP)net system production/(t·km-2) 6326.414 5485.947 7924.189 0.416 0.554总初级生产力/总生物量(TPP/TB)total primary production/total biomass 16.676 13.986 14.738 0.487 0.508总生物量total biomass/(t·km-2) 673.279 800.535 986.977 0.988 0.069联结指数(CI)connectance index 0.203 0.209 0.224 0.942 0.154系统杂食指数(SOI)system omnivory index 0.136 0.139 0.172 0.812 0.285循环指数(FCI)finn's cycling index 2.180 3.840 9.120 0.917 0.186平均能流路径(MPL)finn's mean path length 2.555 2.688 3.486 0.855 0.249
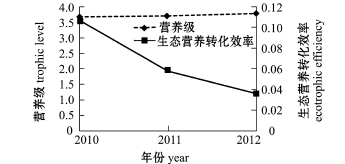
图6 2010—2012年夏季獐子岛人工鱼礁区许氏平鲉估算参数
Fig.6 Parametersofschlegel'srockfish Sebastes schlegelii in Zhangzi Island artificial reef from summer of 2010 to summer of 2012
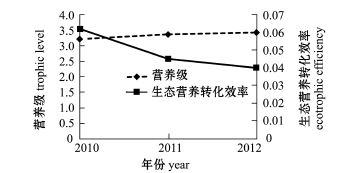
图7 2010—2012年夏季獐子岛人工鱼礁区大泷六线鱼估算参数
Fig.7 Parameters of fatgreenling Hexagrammos otakii in Zhangzi Islandartificial reef from summer of 2010 to summer of 2012
本研究中刺参与海胆的营养级逐渐降低。Pauly等[21]根据联合国粮农组织 (FAO)提供的数据分析表明,1950—1994年全球渔获物的平均营养级从3.3降至2.1,渔获物由高营养级且寿命长的底层鱼类向低营养级且短寿命的中上层鱼类转变。Conover等[22]和Kuparinen等[23]认为,捕捞压力过大可改变鱼类群体的遗传结构,导致个体小且性成熟早的个体更加适应人为选择压力并占据群体的主导地位。獐子岛海域中对刺参与海胆的采捕及底播增殖等人类活动可能影响其种群结构,使种群向低龄化转变,营养级降低。
3.2 营养级的能流特征
模型输出的参数表明,獐子岛海洋牧场人工鱼礁海域生态系统各营养级系统总流量及生物量等参数呈逐级递减趋势,基本符合营养级能量和生物量的金字塔分布特征 (图8)。
第Ⅰ营养级是该系统能量的主要来源,2010—2012年夏季第Ⅰ营养级生物量均逐渐增加,碎屑食物链流通量增大 (图3~图5),初级生产者和碎屑能量转化效率逐渐提升并高于平均能量转化效率12%。陈勇等[24]认为,投放鱼礁形成的上升流将海底深层的营养盐输送至光照充足的上层,促进了浮游动植物的繁殖,提高了海洋初级生产力,碎屑食物链提高了能量利用效率。碎屑流通量的增加及转化效率的提高,加快了有机碎屑在生态系统中的循环速度,对该海域底质环境的改善起到了积极的作用。
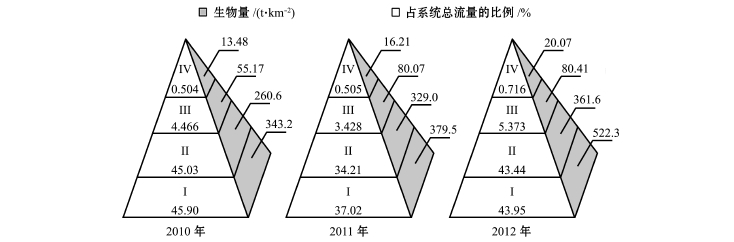
图8 2010—2012年夏季獐子岛人工鱼礁区生态系统总流量与生物量在营养级间的分布
Fig.8 Distribution of throughput and biomass at effective trophic levels in Zhangzi Island artificial reef ecosystem from summer of 2010 to summer of 2012
3.3 功能组生态重叠度的变化
通过分析生态重叠度指数平均值的变化 (表5)可以看出,在该海域皱纹盘鲍的生物量增高的情况下 (表2),其猎物重叠度指数同时升高,反映了皱纹盘鲍在生物量连续增高的情况下发生种内饵料竞争的可能性增加,而该海域皱纹盘鲍捕食重叠度指数升高,则反映了以皱纹盘鲍为食的功能组
参考文献:个数增加,说明皱纹盘鲍在该生态系统内生物量增高的同时,皱纹盘鲍的被食几率也随之增加。林开敏等[25]认为,由于影响生物生长发育的生态因子众多,单个或几个因子的生态位重叠度不能完全反映竞争程度。李契等[15]认为,生态位重叠是利用性竞争的必要条件之一,即生态位重叠度可以反映竞争风险大小。该海域许氏平鲉猎物重叠度指数平均值逐渐升高 (表5),通过生物量调查表明,2012年许氏平鲉的生物量是2010年的3.285倍(表2)。许氏平鲉属北方冷温性底层岩礁性鱼类,是北方近海人工鱼礁区的主要增殖养护对象之一[26-28],该海域许氏平鲉的生物量与生态位重叠度指数的变化规律说明,通过人工鱼礁的投放,在对该鱼类产生较好的诱集与养护效果的同时,增加了其被食几率。
3.4 生态系统总体的特征变化
獐子岛海洋牧场人工鱼礁海域生态系统特征参数 (表6)表明,该生态系统3个时期的总消耗量、总输出量、流向碎屑总量逐年增高,系统规模总体呈增加趋势。从系统功能方面来看,一般情况下,成熟的系统TPP/TR值接近1,NSP值趋近于0。而本研究中3个时期生态系统的TPP/TR值均大于1,NSP值大于0且变化不稳定,说明系统内能量尚有剩余。CI和SOI是表征生态系统内部种群之间复杂程度的指数,越是成熟的系统,复杂程度就越高[29],生态系统的CI和SOI值逐渐升高,说明该生态系统内部稳定性逐渐增强。FCI是系统生产力供给的物质和能量进入再循环的比例,当FCI=0~0.1时,系统属于低再循环率,系统处于发育早期;当FCI>0.5时,说明系统处于高再循环率的成熟期[30]。本研究中3个时期的FCI值均大于0.5,说明该海域物质和能量的再循环率较高,系统成熟度较高。MPL为营养在食物链中流动的长度[31],在成熟的生态系统中,营养流经过的食物链较长[32]。本研究中3个时期的MPL值逐渐增大,说明该生态系统的成熟度有增高的趋势。
综上所述,2010—2012年獐子岛人工鱼礁海区生态系统规模、能流路径长度、能流循环速率和内部联系程度逐渐升高,稳定性逐渐增强,说明该系统成熟度较高,有较高的系统净生产力。模型计算结果表明,个别海珍品存在营养级降低的情况,建议在系统成熟过程中对该海域内生物的采捕规格进行合理控制,如适当施行降低捕捞强度或提高开捕规格等手段,保证人工鱼礁区资源的可持续发展。
3.5 模型质量评价
Morissette等[33]对世界范围内150个 Ecopath模型的输入参数进行模型质量分析,得出Pedigree指数范围为0.16~0.68。本研究中,2010—2012 年3个时期的Ecopath模型Pedigree指数分别为0.554、0.619和0.567,质量相对较好,可信度较高,但仍可以通过细化功能组的分类以及提高数据采集精度等方法提高模型质量。相对于自然海域生态系统,人工鱼礁海域生态系统的形成和发展受人类活动影响较大,应加大对模型所需数据的采集密度与时间跨度,力求更加准确、客观和真实地反映该生态系统特征的变化。
[1] 田涛,陈勇,陈辰,等.獐子岛海洋牧场海域人工鱼礁区投礁前的生态环境调查与评估[J].大连海洋大学学报,2014,29(1):75-81.
[2] 张继红,方建光,王诗欢.大连獐子岛海域虾夷扇贝养殖容量[J].水产学报,2008,32(2):236-241.
[3] 于莹,张武昌,张光涛,等.2010年两个航次獐子岛海域浮游纤毛虫丰度和生物量[J].生态学报,2012,32(22):7220-7229.
[4] 尹洁慧.獐子岛海域浮游动物群落时空变化特征及其与环境因子之间的关系[D].北京:中国科学院大学,2013.
[5] 王全超,韩庆喜,李宝泉.辽宁獐子岛马牙滩潮间带及近岸海区大型底栖动物群落特征[J].生物多样性,2013,21(1):11-18.
[6] 陈勇,杨军,田涛,等.獐子岛海洋牧场人工鱼礁区鱼类资源养护效果的初步研究[J].大连海洋大学学报,2014,29(2):183-187.
[7] 仝龄.Ecopath——一种生态系统能量平衡评估模式[J].海洋水产研究,1999,20(2):102-107.
[8] 李永刚,汪振华,章守宇.嵊泗人工鱼礁海区生态系统能量流动模型初探[J].海洋渔业,2007,29(3):226-234.
[9] 赵静,章守宇,许敏.枸杞海藻场生态系统能量流动模型初探[J].上海海洋大学学报,2010,19(1):98-104.
[10] 吴忠鑫,张秀梅,张磊,等.基于Ecopath模型的荣成俚岛人工鱼礁区生态系统结构和功能评价[J].应用生态学报,2012,23(10):2878-2886.
[11] Su Jilan,Tang Qisheng.Study on ecosystem dynamics in coastal ocean:II.processes of the Bohai Sea ecosystem dynamics[M]. Beijing:China Science Press,2002.
[12] Pauly D,Soriano-Bartza M L,Palomares M L D,et al.Improved construction,parameterization,and interpretation of steady-state ecosystem models[C]//Christensen V,Pauly D.Trophic Models of Aquatic Ecosystems.Manila:ICLARM,1993.
[13] 仝龄,唐启升,Daniel P.渤海生态通道模型初探[J].应用生态学报,2000,11(3):435-440.
[14] Okey T A,Banks S,Born A F,et al.A trophic model of a Galápagos subtidal rocky reef for evaluating fisheries and conservation strategies[J].Ecological Modelling,2004,172(2-4):383-401.
[15] 李契,朱金兆,朱清科.生态位理论及其测度研究进展[J].北京林业大学学报,2003,25(1):100-107.
[16] Hurlbert S H.The measurement of niche overlap and some relatives[J].Ecology,1978,59(1):67-77.
[17] Pianka E R.The structure of lizard communities[J].Annual Review of Ecology and Systematics,1973,4:53-74.
[18] Baird D,Asmus H,Asmus R.Energy flow of a boreal intertidal ecosystem,the Sylt-RØmØ Bight[J].Marine Ecology Progress Series,2004,279:45-61.
[19] 唐启升,苏纪兰,孙松,等.中国近海生态系统动力学研究进展[J].地球科学进展,2005,20(12):1288-1299.
[20] 张波,唐启升.渤、黄、东海高营养层次重要生物资源种类的营养级研究[J].海洋科学进展,2004,22(4):393-404.
[21] Pauly D,Christensen V.Primary production required to sustain global fisheries[J].Nature,1994,374:255-257.
[22] Conover D O,Munch S B.Sustaining fisheries yields over evolutionary time scales[J].Science,2002,297:94-96.
[23] Kuparinen A,Merilä J.Detecting and managing fisheries-induced evolution[J].Trends in Ecology&Evolution,2007,22(12):652-659.
[24] 陈勇,于长清,张国胜,等.人工鱼礁的环境功能与集鱼效果[J].大连水产学院学报,2002,17(1):64-69.
[25] 林开敏,郭玉硕.生态位理论及其应用研究进展[J].福建林学院学报,2001,21(3):283-287.
[26] 童玉和,郭学武.两种岩礁鱼类的食物竞争实验[J].中国水产科学,2009,16(4):541-549.
[27] 张硕,孙满昌,陈勇.人工鱼礁模型对大泷六线鱼和许氏平鲉幼鱼个体的诱集效果[J].大连水产学院学报,2008,23(1):13-19.
[28] 冯昭信,韩华.大泷六线鱼资源合理利用的研究[J].大连水产学院学报,1998,13(2):24-28.
[29] Christensen V,Walters C J,Pauly D,et al.Ecopath with Ecosim version 6 user guide[M].Vancouver:University of British Columbia,2008.
[30] 林群,金显仕,郭学武,等.基于Ecopath模型的长江口及毗邻水域生态系统结构和能量流动研究[J].水生态学杂志,2009,2(2):28-36.
[31] Odum E P.The strategy of ecosystem development[J].Science,1969,164:262-270.
[32] Christensen V,Walters C J,Pauly D.Ecopath with Ecosim:a user's guide[M].Vancouver:Fisheries Centre,University of British Columbia,2005:154.
[33] Morissette L,Hammill M O,Savenkoff C.The trophic role of marine mammals in the northern Gulf of St.Lawrence[J].Marine Mammal Science,2006,22(1):74-103.
Abstract:An Ecopath model of artificial reef ecosystem was established in the Zhangzi Island by the software known as Ecopath with Ecosim(EwE)6.4 and the energy flow pattern and the system attributes were evaluated in the ecosystem in summer from 2010 to 2012 in order to investigate the structure and function of Zhangzi Island artificial reef ecosystem.The model was found to be comprised of 23 functional groups,including the main processes of the energy flow in the artificial reef ecosystem in Zhangzi Island.The results showed that yesso scallop Patinopecten yessoensis,Schlegel's rockfish Sebastes schlegelii,fat greenling Hexagrammos otakii and other marine organisms had higher trophic level value from summer in 2010 to summer in 2012.The prey overlap index was found to be increased from 0.123 to 0.139 in disk abalone Haliotis discus hannai and the prey overlap index from 0.112 to 0.165 in disk abalone Haliotis discus hannai.The predator overlap index was increased from 0.063 to 0.185 in Schlegel's rockfish and from 0.062 to 0.132 in fat greenling.The ecological efficiency(EE)index was increased from 11.1% in summer of 2010 to 14.4%in summer of 2012,with finn's cycling index(FCI)of over 0.1 and high net system production(NSP).The system omnivory index(SOI)was increased from 0.136 to 0.172,the connectance index (CI)from 0.203 to 0.224,and Finn's mean path length(MPL)from 2.555 to 3.486 in Zhangzi Island artificial reef ecosystem in summer from 2010 to 2012.In conclusion,the Zhangzi Island artificial reef ecosystem was found to be in stable mature stage from 2010 to 2012.
Key words:artificial reef;ecosystem structure;energy flow;niche overlap index;ecopath model
Environmental suitability of sea cucumber Apostichopus japonicus based on a habitat suitability index model
LI Lin-han,CHEN Yong,TIAN Tao,LIU Yong-hu,YU Ya-qun,ZHANG Hao-yu
(Center for Marine Ranching Engineering Science Research of Liaoning,Dalian Ocean University,Dalian 116023,China)
Abstract:Suitability of environment was studied in sea cucumber Apostichopus japonicus based on a habitat suitability index(HSI)model.The suitability index(SI)value and HSI values of sea cucumber in Dachangshan waters,temperature,salinity,dissolved oxygen,sediment environment and percentage content of sediment sand and gravel as factors,were estimated by experience value assignment method,mathematical statistics method,and geometric average method.It was found that the average of HSI values were 0.66,0.59,0.67,0.46,0.66,0.65 and 0.65 in 7 voyages from 2013 to 2014,significantly lower in winter(HSI=0.46)than in other seasons,and relatively high suitability in spring(HSI=0.66)and autumn(HSI=0.67,HSI=0.65).In the same voyage,November 2014,for example,the HSI value at each station was 0.66,0.67,0.65,0.62,0.63,0.62 and 0.67,with high suitability at DCS01,DCS02 and DCS07 positions.It is suggested by the calculation that sea cucumber be released at DCS01,DCS02 and DCS07 in the spring and autumn.
Key words:Apostichopus japonicas;habitat suitability index;environment suitability
Structure and function of an artificial reef ecosystem in Zhangzi Island based on Ecopath model
XU Zhen-xing,CHEN Yong,TIAN Tao,LIU Yong-hu,YIN Zeng-qiang,LIU Han-chao
(Center for Marine Ranching Engineering Science Research of Liaoning,Dalian Ocean University,Dalian 116023,China)
中图分类号:S932.8
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2016.01.014
文章编号:2095-1388(2016)01-0080-05
收稿日期:2015-05-25
基金项目:国家海洋公益性行业科研专项 (201205023);国家 “十二五”科技支撑计划项目 (2012BAD18B03,2013BAD23B02)
作者简介:李林晗 (1989—),女,硕士研究生。E-mail:llh0131@sina.com
通信作者:田涛 (1979—),男,博士,副教授。E-mail:ttbeyond@126.com
文献标志码:
中图分类号:S932.2A