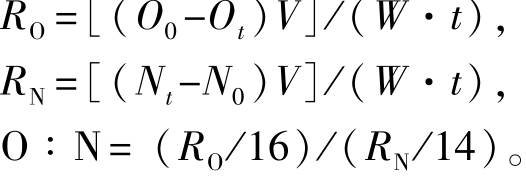
摘要:为了研究不同硒源对刺参Apostichopus japonicus特定生长率和呼吸代谢指标的影响,以刺参幼参和成参为研究对象,分别强化投喂硒添加量为0.6 mg/kg的硒代蛋氨酸、硒酸钠和亚硒酸钠饲料,试验周期为60 d。结果表明:硒添加组成参和幼参的特定生长率均显著大于对照组 (P<0.05),其中硒代蛋氨酸组最大,且显著大于硒酸钠组和亚硒酸钠组 (P<0.05),除硒代蛋氨酸组外,其他各试验组成参的特定生长率均大于幼参;硒添加组成参和幼参的耗氧率均显著大于对照组 (P<0.05),其中硒代蛋氨酸组最小,显著小于硒酸钠组和亚硒酸钠组 (P<0.05),对应组中,幼参耗氧率大于成参;硒添加组成参和幼参的排氨率均低于对照组,除硒代蛋氨酸组成参显著低于对照组 (P<0.05)外,其余硒添加组与对照组均无显著性差异 (P>0.05),硒代蛋氨酸组的排氨率最小,对应组中,幼参排氨率大于成参;硒添加组成参和幼参的O/N值均显著大于对照组 (P<0.05),其中硒代蛋氨酸组最小,但硒添加组间无显著性差异 (P>0.05),对应组中,成参的O/N值大于幼参。研究表明,饲料中添加硒能够提高刺参的代谢率,促进刺参生长,其中有机硒 (硒代蛋氨酸硒)的促生长效果好于无机硒 (硒酸钠和亚硒酸钠),且硒代蛋氨酸对幼参的促生长效果要好于成参。
关键词:刺参;硒源;特定生长率;耗氧率;排氨率;氧氮比
硒源选择是富硒农水产品开发中的关键技术[1],硒源一般包括两大类,一类是以硒代蛋氨酸为代表的有机硒,另一类是以硒酸钠和亚硒酸钠为代表的无机硒。通常无机硒中硒含量较高,但生物利用率较低,而有机硒生物利用率较高,但生产工艺复杂,成本较高[2-3]。不同硒源对不同动物生理代谢过程的影响不尽相同。张华[4]在对兔免疫功能的研究中发现,通过10 d的硒强化,无机硒饲料组兔出血症抗体较有机硒组提高了28.1%;魏文志等[5]和杨丽坤等[6]在对有机硒与无机硒的比较研究中发现,有机硒组异育银鲫的特定生长率最大提高了14.59%,有机硒组中华米虾的孵化率最大提高了23.8%;王吉桥等[1]在对刺参 Apostichopus japonicus幼参的研究中发现,蛋氨酸硒强化组的SOD和ACP活性最强,较亚硒酸钠饲料组分别提高了12.5%和51.2%。刺参作为中国主要的水产养殖种类之一,在刺参的富硒强化培养过程中,硒源的选择显得尤为重要。
呼吸代谢是生物能量学研究的重要内容,在相关研究中,人们经常利用呼吸代谢指标评价动物对营养元素的利用效果。赵元凤等[7]在研究螯合盐作为营养添加剂的效果时发现,螯合盐组罗非鱼耗氧率在降低27.4%的条件下,鱼体蛋白质和脂肪的比值提高了15.8%,饵料系数降低了33.4%;闫海亮[8]和张苏江[9]在对锌和铜元素的营养效果研究时,通过对呼吸代谢指标观察发现,獭兔饲料中锌的添加量为40~120 mg/kg,猪饲料中铜添加量为100~300 mg/kg时,随着锌、铜营养元素含量的增加,尿素氮浓度逐渐降低,最多降低了10.6%和9.1%,同时机体总蛋白质含量逐渐增加,最大增加量为1.3%和9.2%。目前,有关刺参呼吸代谢研究中,温度、盐度、摄食和体质量等因素影响的相关研究报道较多[10-14],但微量元素对刺参呼吸代谢影响的相关研究尚未见报道。针对海参富硒技术,作者先后围绕相关理论和技术进行了一些研究与探讨[15-16],本研究中通过硒代蛋氨酸、亚硒酸钠、硒酸钠3种硒化合物对刺参进行强化投喂,研究不同硒源对刺参呼吸代谢的影响,从营养生理角度深入探析硒在刺参强化投喂过程中的作用机理,旨在为相关技术研发提供理论依据。
1.1 材料
试验用刺参幼参360头,初始体质量为(2.28±0.26)g,成参 180头,初始体质量为(54.97±9.49)g,分别在试验条件下驯养10 d。
基础饲料为市售海参配合粉末饲料,含粗蛋白质为 15.00%,粗脂肪为 3.21%,粗灰分为47.33%,硒含量为0.2981 mg/kg,在60℃下烘干至恒重备用。
试验仪器主要有网箱 (100 cm×100 cm×100 cm)、蓝色波纹板 (90 cm×90 cm)、YSI ProPlus型手持式野外/实验室两用测量仪、分析天平 (精度为0.01 g)、电子天平 (精度为0.05 g)、721分光光度计等。
试验药品硒代蛋氨酸、亚硒酸钠和硒酸钠均为分析纯,由成都格雷西亚化学技术有限公司生产;硫代硫酸钠、碘化钾、氯化锰、氢氧化钠、硫酸、硫酸铵、盐酸奈乙二胺均为分析纯,硝酸、高氯酸、盐酸均为优级纯,可溶性淀粉为分析纯。
1.2 方法
1.2.1 试验饲料的制备 准确称取15 g淀粉4份,分别倒入375 mL水中制成淀粉液,再将硒添加量为0.6 mg/kg(饲料)的硒代蛋氨酸、硒酸钠、亚硒酸钠分别溶解到其中3份对应的淀粉液中,制成硒代蛋氨酸组、硒酸钠组、亚硒酸钠组3组饲料原料,并将不添加硒的饲料设为对照组。加热至80℃使其糊化,再准确称取500 g市售配合饲料4份,分别与上述4组原料混合均匀,用绞肉机压粒制成4组试验饲料,在烤箱 (60℃)中烘干至恒重备用。
1.2.2 试验设计 将驯养后的幼参和成参分别按对照组、硒代蛋氨酸组、硒酸钠组、硒酸钠组随机平均分成4组,每组设3个平行,幼参组每个平行放幼参30头,成参组每个平行放成参15头,将分好组的刺参分别放入内置有蓝色波纹板的网箱中饲养,分别投喂对应的饲料。
试验在刺参养殖池塘中进行,自然光照,水温为13℃,盐度为27,pH为8.1。每日17:00投喂试验饲料,投喂量为刺参体质量的3.0% ~5.0%,并随刺参生长和摄食情况适当增减,试验周期为60 d,每20 d倒箱一次。
1.2.3 指标的测定与计算
(1)生长指标。将刺参放在筛绢网上静置10 min后,用滤纸吸干其体表水分,用电子天平称量其质量。计算公式为
存活率=nt/n0×100%,
特定生长率=(ln Wt-ln W0)/t×100%。
其中:n0、nt分别为试验初和试验末刺参的存活数;W0和Wt分别为试验初和试验末刺参的体质量(g);t为饲养时间 (d)。
(2)耗氧率和排氨率。试验结束时,将两种规格的刺参饥饿72 h,采用静水法测定试验组刺参的耗氧率和排氨率。幼参和成参按试验分组测定,每组设3个平行,幼参组从每个平行随机取10头,成参组从每个平行随机取1头。使用容积为2.5 L的呼吸瓶,同时设空白对照组 (不放刺参),用海水贮满呼吸瓶,呼吸瓶密闭5 h后,采用次溴酸钠氧化法和Winkler碘量法[17]测定水样中的溶解氧和总氨氮浓度,依据下式计算耗氧率[RO,mg/(g· h)]、排氨率[RN,μg/(g·h)]和氧氮比:
其中:O0和Ot分别为试验结束时对照组、试验组水体中溶解氧质量浓度 (mg/L);N0和Nt分别为试验结束时对照组、试验组水体中总氨氮质量浓度(mg/L);t为试验持续时间 (h);W为刺参的湿质量 (g);V为试验水体体积 (L)。
1.3 数据处理
试验数据均以平均值±标准差表示,采用SPSS 16.0软件进行单因素方差分析,采用LSD法进行多重比较,显著性水平设为0.05。
2.1 幼参和成参的存活率和特定生长率
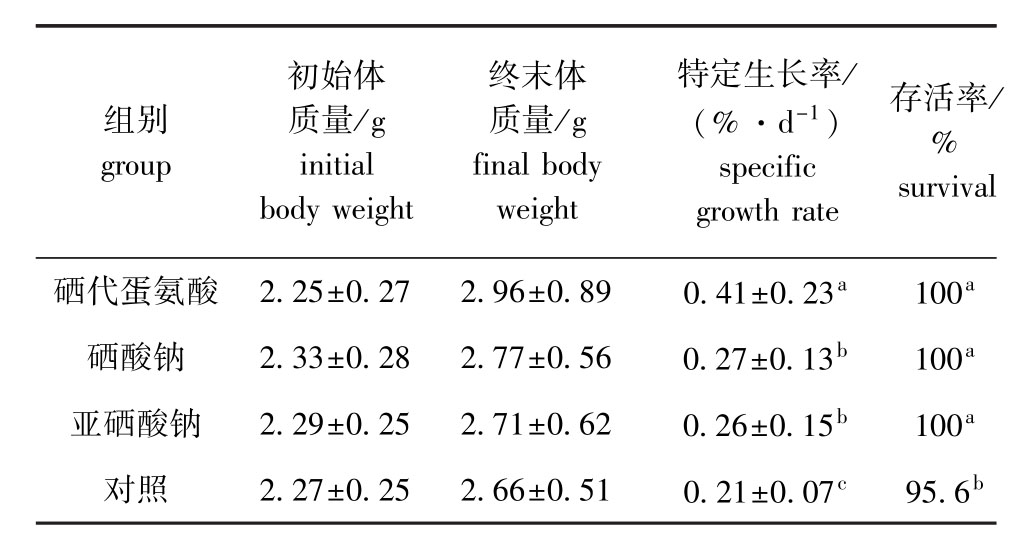
从表1可见:在整个试验阶段,硒添加组幼参的存活率均为100%,对照组为95.6%,硒添加组刺参的存活率均显著高于对照组 (P<0.05);幼参各组的特定生长率则表现为硒添加组均显著高于对照组 (P<0.05),而硒添加组中硒代蛋氨酸组的特定生长率最高,亚硒酸钠组的特定生长率最低,硒代蛋氨酸组显著高于硒酸钠组和亚硒酸钠组 (P<0.05),而硒酸钠组与亚硒酸钠组间无显著性差异(P>0.05)。
表1 各组幼参的特定生长率和存活率
Tab.1 Growth and survival of the juvenile sea cucumber in different groups
注:同列中标有不同小写字母者表示组间有显著性差异 (P<0.05),标有相同小写字母者表示组间无显著性差异 (P>0.05),下同
Note:The means with different letters within the same column are significant differences at the 0.05 probability level,and the means with the same letters within the same column are not significant differences,et sequentia
组别group初始体质量/g initial body weight终末体质量/g final body weight特定生长率/ (%·d-1)specific growth rate存活率% survival硒代蛋氨酸 2.25±0.27 2.96±0.89 0.41±0.23a 100 / a硒酸钠 2.33±0.28 2.77±0.56 0.27±0.13b 100a亚硒酸钠 2.29±0.25 2.71±0.62 0.26±0.15b 100a对照 2.27±0.25 2.66±0.51 0.21±0.07c 95.6b
从表2可见:在整个试验阶段,成参各组的存活率均为100%;成参各组的特定生长率则表现为硒添加组均显著高于对照组 (P<0.05),而硒添加组中硒代蛋氨酸组的特定生长率最高,硒酸钠组最低,硒代蛋氨酸组显著高于硒酸钠组和亚硒酸钠组(P<0.05),而硒酸钠组和亚硒酸钠组间无显著性差异 (P>0.05)。与表1相比,除硒代蛋氨酸组外,其他各组中成参的特定生长率均大于幼参。/
表2 各组成参的特定生长率和存活率
Tab.2 Growth and survival of the adult sea cucumber in different groups
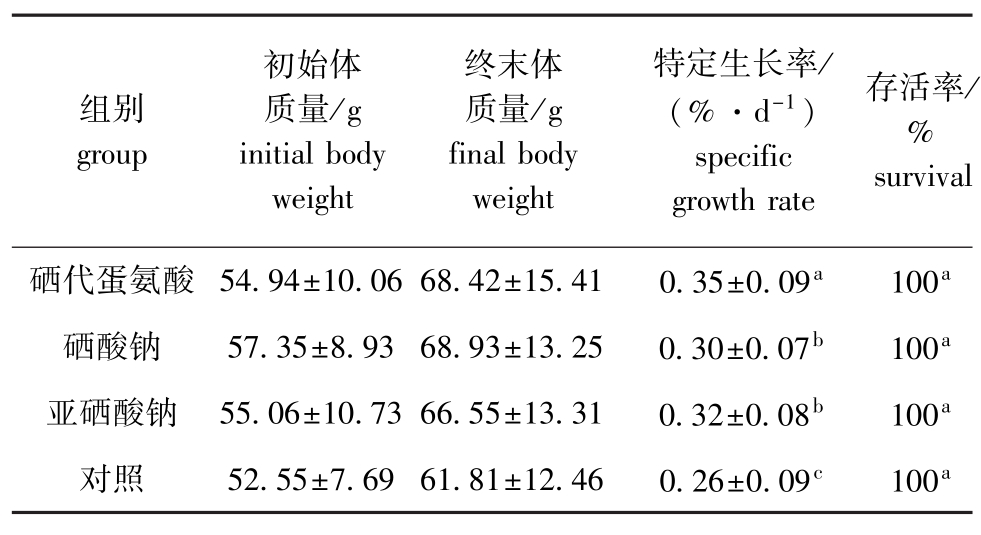
组别group初始体质量/g initial body weight终末体质量/g final body weight特定生长率/ (%·d-1)specific growth rate存活率% survival硒代蛋氨酸 54.94±10.06 68.42±15.41 0.35±0.09a 100a硒酸钠 57.35±8.93 68.93±13.25 0.30±0.07b 100a亚硒酸钠 55.06±10.73 66.55±13.31 0.32±0.08b 100a对照 52.55±7.69 61.81±12.46 0.26±0.09c 100a
2.2 幼参和成参的耗氧率和排氨率
从图1可见:经60 d硒饲料强化投喂后,幼参硒添加组的耗氧率较对照组均有显著提高 (P<0.05),其中亚硒酸钠组的耗氧率最大,为20.32 mg/(g·h),较对照组提高了33.9%,硒代蛋氨酸组最小,为18.75 mg/(g·h),较对照组提高了23.5%;成参硒添加组刺参的耗氧率也较对照组均有显著提高 (P<0.05),其中亚硒酸钠组的耗氧率最大,为 17.25 mg/(g·h),较对照组提高了41.5%,硒代蛋氨酸组最小, 为 15.57 mg/(g·h),较对照组提高了27.7%。在相应的各试验组中,幼参的耗氧率均大于成参。
从图1还可见:经60 d硒饲料强化投喂后,幼参硒添加组排氨率较对照组均有所下降,但无显著性差异 (P>0.05),硒添加组中硒代蛋氨酸组的排氨率最小,为1.49 μg/(g·h),较对照组下降了5.1%,亚硒酸钠组最大,为1.56 μg/(g·h),较对照组下降了1.0%;成参硒添加组的排氨率较对照组均有所下降,仅硒代蛋氨酸组较对照组显著下降 (P<0.05),硒添加组中硒代蛋氨酸组的排氨率最小,为0.96 μg/(g·h),较对照组下降了15.0%,亚硒酸钠组和硒酸钠组,均为 1.02 μg/(g·h),较对照组下降了9.7%。在相应各试验组中,幼参的排氨率均大于成参。
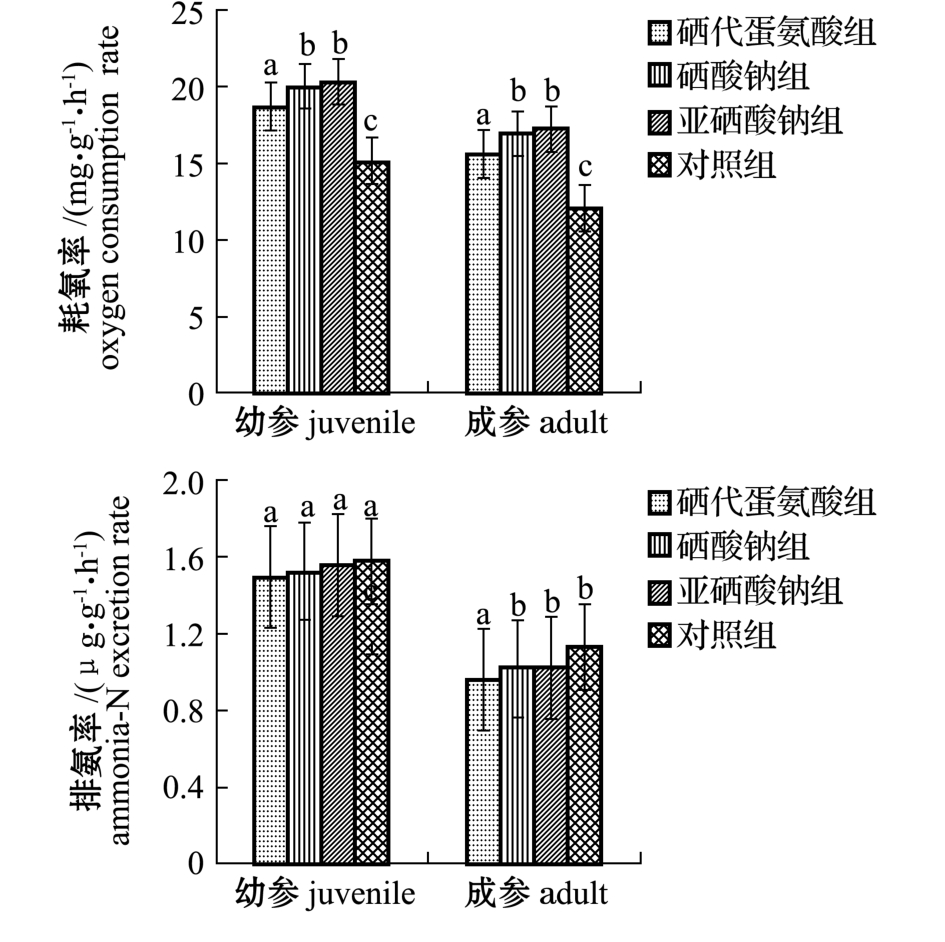
图1 各组刺参的耗氧率和排氨率
Fig.1 Oxygen consumption rate and ammonia-N excretion rate of sea cucumber in different groups
注:标有不同小写字母者表示组间有显著性差异 (P<0.05),标有相同小写字母者表示组间无显著性差异 (P>0.05)
Note:The means with different letters are significant differences at the 0.05 probability level,and the means with the same letters are not significant differences
从表3可见:幼参硒添加组O/N值均显著大于对照组 (P<0.05),其中硒酸钠组最大,较对照组提高了36.4%,硒代蛋氨酸组最小,较对照组提高了29.8%;成参硒添加组的O/N值均显著大于对照组 (P<0.05),其中亚硒酸钠组最大,较对照组提高了56.9%,硒代蛋氨酸组最小,较对照组提高了51.1%。在相应各试验组中,成参的O/N值均大于幼参。
表3 各组刺参的O/N值
Tab.3 O/N ratio of sea cucumber fed the diets containing various types of selenium
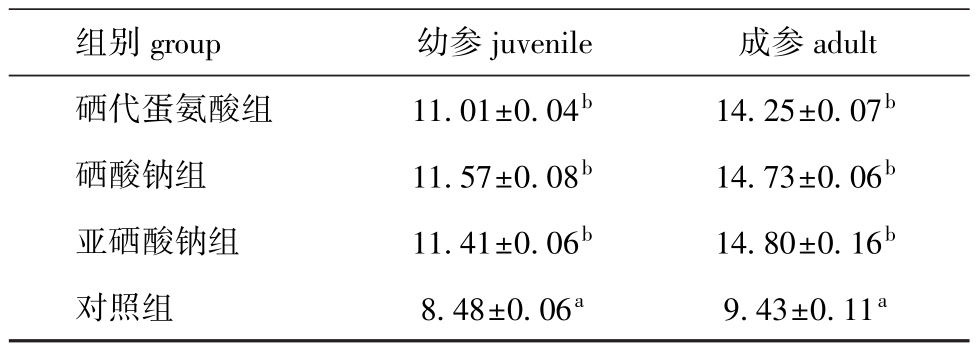
组别group 幼参juvenile 成参adult硒代蛋氨酸组 11.01±0.04b 14.25±0.07b硒酸钠组 11.57±0.08b 14.73±0.06b亚硒酸钠组 11.41±0.06b 14.80±0.16b对照组 8.48±0.06a 9.43±0.11a
3.1 不同硒源对刺参特定生长率的影响
硒是生物体必需的微量营养元素,缺硒会导致生物体生理机能下降,抑制机体生长[18]。
研究表明,在日粮饲料中添加适量的硒能够提高机体谷胱甘肽过氧化物酶 (GSH-Px)、蛋白酶、淀粉酶和脂肪酶的活性,进而提高饲养对象的特定生长率[5,19-21]。本研究中,刺参硒强化试验组与对照组相比均表现为特定生长率提高,最大提高了95.2%。这与对虹鳟[22]、中国对虾[23]、 异育银鲫[5,24]等许多水产动物的硒强化研究结果一致。由此可见,硒对刺参的生长具有促进作用,在日粮中添加适量的硒是有必要的。在硒的选择研究方面,郑宗林等[25]通过对叉尾鮰的研究认为,有机硒(硒代蛋氨酸和酵母硒)对生长的促进作用比无机硒 (亚硒酸钠)提高了269% ~336%。本研究中,硒代蛋氨酸较硒酸钠和亚硒酸钠的促生长效果差距不大,刺参的特定生长率最大差距仅为57.7%。在无机硒的选择方面,有关亚硒酸钠作为水产动物硒源的研究较多,对硒酸钠的研究报道较少[1-6],本研究中,成参组亚硒酸钠较硒酸钠的促生长效果提高6.7%,二者无显著性差异 (P>0.05),幼参组硒酸钠的促生长效果好于亚硒酸钠,前者较后者提高3.8%,二者也无显著性差异 (P>0.05),这表明以硒酸钠作为刺参硒源可达到与亚硒酸钠相同的促生长效果。从幼参和成参的特定生长率进行比较分析,无机硒组 (硒酸钠、亚硒酸钠)和对照组的特定生长率均为成参大于幼参,其中硒酸钠组成参的特定生长率高出幼参11.1%,亚硒酸钠组高出23.1%,对照组高出23.8%。梁其正等[26]在相关研究中对成体增重率快于幼体的现象解释为,成体对饵料的摄食能力和消化能力较强。值得注意的是,本研究中,硒代蛋氨酸组幼参与成参的特定生长率表现出的差异正好相反,幼参较成参高出17.1%,上述个体规格在生长指标方面表现出的差异与硒源的相关性目前尚未见报道,有待于进一步研究。
3.2 不同硒源对刺参耗氧率和排氨率的影响
大量研究表明,适量提高饲料中的营养水平,可以提高机体的呼吸和代谢水平[27-30],同时日粮中营养成分与机体的氨氮代谢呈显著的相关性[31]。本研究中,硒强化投喂的各试验组刺参的耗氧率都有显著提高,较对照组最大提高了41.5%,根据张硕等[32]、 孙侦龙等[29]、 李宝泉等[33]、 袁秀堂等[13]、包杰等[11]在相关研究中的观点,刺参耗氧率的提高表明,硒作为微量营养元素,适量添加能够提高饲料整体的营养水平。本研究中,试验各组刺参排氨率较对照组均表现出降低,最大降低了15.0%,根据王桂芹等[30]在相关研究中的观点,刺参排氨率的降低表明,硒作为微量营养元素,适量添加能够提高饲料中蛋白质的有效利用率。
赵元凤等[34]研究认为,以螯合微量元素作为鱼类、贝类营养添加剂,可使耗氧率较无机盐组降低39.5%。Cheng等[35]研究认为,通过大豆蛋白补充赖氨酸,可降低虹鳟排氨率,从此角度分析,本研究在硒源的比较上,有机硒组 (硒代蛋氨酸)刺参耗氧率最低,且显著低于无机硒组 (亚硒酸钠、硒酸钠) (P<0.05),最大降低了9.7%;有机硒组刺参排氨率也是最低,且显著低于无机硒组(P<0.05),最大降低了5.9%。综上所述,从能量代谢角度比较,有机硒较无机硒更能通过增大生长能和排泄能的比例,降低代谢能比例的方式,提高饲料营养效率[36]。
相关研究表明,单位个体的耗氧率和排氨率会随着体质量的增加而下降[9,37-40]。本研究中,相同饲料组刺参的耗氧率和排氨率均随规格的增大而减小,成参的耗氧率较幼参最大降低了19.7%,排氨率最大降低了35.6%,与不同规格海胆[37]、九孔鲍[38]、黑脊倒刺鲃[39]等的呼吸代谢规律一致。
3.3 不同硒源对刺参O/N值的影响
氧氮比 (O/N)是表示动物呼吸底物的重要参数,海洋动物对能量的需求和对营养物质的利用情况可以用氧氮比进行评估[11]。理论上,O/N<10时,生物以蛋白质代谢为主;O/N>10时,则主要以脂肪和碳水化合物代谢为主[33]。本试验条件下,对照组刺参O/N值均小于10(8.48~9.43),说明其代谢能量主要由蛋白质提供;而硒添加组O/N值均大于10(11.01~14.80),说明其主要代谢能量由脂肪和碳水化合物提供。由此推断,在饲料中添加一定水平的硒,有助于提高刺参饲料中脂肪和碳水化合物的利用效果,对于改善刺参的营养结构和提高蛋白质的沉积效果具有重要意义。本研究中,有机硒组的O/N值均小于无机硒组,根据对刺参[10,13,33]、 对虾[32]、 栉孔扇贝[41]等水产动物O/N值受培育环境影响的研究结果分析,可能与有机硒和无机硒在刺参机体内的代谢途径不同有关,具体原因有待进一步研究。本研究中对应的饲料组中,成参的O/N值均高于幼参,最大值可达29.7%,说明在相同的试验条件下,成参蛋白质的沉积效果好于幼参,这与对大黄鱼[31]、栉孔扇贝[41]、波纹龙虾[42]等水产动物的研究结果一致。
从营养生理角度来看,从食物中摄取的蛋白质、脂肪和糖经机体消化吸收后,一部分用作能量消耗,一部分用作生长,从总量平衡分析,在摄入量一定的条件下,蛋白质作为能量消耗的部分越少,作为自身积累的部分就越多,同时该饲料各营养成分的配比就越合理,饲料营养价值就越高[43]。本研究结果表明,在基础饲料中适量添加硒能够优化饲料营养结构,提高刺参代谢率,降低能耗,进而促进刺参生长。本试验中,硒代蛋氨酸组刺参的耗氧率和排氨率均最小,但该组刺参的特定生长率均最大,说明硒代蛋氨酸在促刺参生长上效果最好,并且就促生长而言,硒代蛋氨酸在幼参中的效果好于成参。由此可知,在日粮中加入硒代蛋氨酸对刺参的生长效果最好,尤其对幼参,鉴于对无机硒的研究效果,并综合考虑原料成本和采购渠道等因素,也可灵活选择亚硒酸钠和硒酸钠作为硒源。
参考文献:
[1] 王吉桥,王志香,于红艳,等.饲料中不同类型的硒对仿刺参幼参生长和免疫指标的影响[J].大连海洋大学学报,2011,26 (4):306-311.
[2] 金丰秋,金其荣.硒酵母(有机硒)与硒酸、亚硒酸(无机硒)在安全性方面的差别[J].应用技术,2001(11):33-34.
[3] 胡先勤,刘立鹤,陈见,等.酵母硒在鲫鱼饲料中的应用[J].中国牧业通讯,2010(1):33-34.
[4] 张华.不同硒源对兔免疫功能和抗氧化能力影响的研究[D].南京:南京农业大学,2004.
[5] 魏文志,杨志强,罗方妮,等.饲料中添加有机硒对异育银鲫生长的影响[J].淡水渔业,2001,31(3):45-46.
[6] 杨丽坤,蔡端波,李泽健,等.硒和酵母对中华米虾孵化率的影响[J].安徽农业科学,2011,39(6):3666,3748.
[7] 赵元凤,吕景才,郭庆,等.氨基酸螯合盐对罗非鱼促生长作用的研究[J].中国水产科学,1997,4(6):40-43.
[8] 闫海亮.日粮蛋氨酸锌对獭兔生产性能、生化指标的影响[D].长春:吉林大学,2008.
[9] 张苏江.日粮铜水平对生长猪生长性能及生化指标影响的研究[D].长春:中国人民解放军军需大学,2001.
[10] 隋佳佳,董双林,田相利,等.光谱和体重对刺参耗氧率和排氨率的影响[J].中国海洋大学学报,2010,40(3):61-64.
[11] 包杰,姜宏波,董双林,等.红刺参和青刺参耗氧率与排氨率的比较研究[J].水产学报,2013,37(11):1689-1696.
[12] 周玮,桑田成,苏延明,等.饥饿对仿刺参苗种生长及呼吸代谢的影响[J].大连水产学院学报,2009,24(3):77-80.
[13] 袁秀堂,杨红生,周毅,等.盐度对刺参(Apostichopus japonicus)呼吸和排泄的影响[J].海洋与湖沼,2006,37(4):348-353.
[14] 包杰.环境因子对青刺参和红刺参(Apostichopus japonicus)代谢与生长及其机制的影响[D].青岛:中国海洋大学,2008.
[15] 周玮,刘剑波,张爱丽,等.富硒海参饲料加工与使用方法对硒元素溶失率的影响[J].大连海洋大学学报,2015,30(1):52-55.
[16] 周玮,张津源,王祖峰,等.3种硒化合物对刺参的急性毒性比较[J].大连海洋大学学报,2014,29(6):629-632.
[17] 国家海洋局.GB17378.4-2007海洋监测规范第4部分:海水分析[S].北京:中国标准出版社,2008.
[18] 张联合,郁飞燕,苗艳芳.硒在人和动物健康上的研究[J].安徽农业科学,2007,35(21):6688-6690.
[19] 梁萌青,王家林,常青,等.饲料中硒的添加水平对鲈鱼生长性能及相关酶活性的影响[J].中国水产科学,2006,13(6):1017-1022.
[20] Raymond F,Burk M D.Selenium an antioxidant nutrient[J].Nutrition in Clinical Care,2002,5(2):75-79.
[21] 苏传福.草鱼幼鱼硒的营养需要研究[D].重庆:西南大学,2008.
[22] 张颖,赵吉伟,卢彤岩,等.不同硒源对虹鳟生长性能及抗氧化能力的影响[J].水产学杂志,2005,18(2):28-34.
[23] 王安利,王维娜,刘存岐,等.饲料中硒含量对中国对虾生长及其体内含量的影响[J].水产学报,1994,18(3):245-248.
[24] 华雪铭,周洪琪,邱小琮,等.饲料中添加芽抱杆菌和硒酵母对异育银鲫的生长及抗病力的影响[J].水产学报,2001,25 (5):448-453.
[25] 郑宗林,黄朝芳,汪道好.不同硒源对叉尾鮰生长性能影响的对比试验[J].粮食与饲料工业,2002(3):30-32.
[26] 梁正其,姚俊杰,张红星.投喂鲢鱼肉对不同规格大鲵苗种生长的影响[J].河北渔业,2011(2):21-23.
[27] Mcpherson B F.Feeding and oxygen uptake of the tropical sea urchin Eucidaris trabuloides(Lamarck)[J].Biol Bull,1968,135 (2):308-321.
[28] 孙侦龙.刺参养殖主要营养要素代谢过程的研究[D].青岛:中国海洋大学,2012.
[29] 孙侦龙,高勤峰,董双林,等.不同配比饲料对刺参C、N、P营养盐收支的影响[J].中国海洋大学学报,2012,42(S1):67-74.
[30] 王桂芹,周洪琪,陈建明,等.饲料蛋白对翘嘴鲌氮排泄的影响[J].华南农业大学学报,2008,29(1):92-96.
[31] 沈勤,徐善良,严小军,等.饲料及体重对大黄鱼排氨率影响的初步研究[J].宁波大学学报:理工版,2008,21(3):318-321.
[32] 张硕,董双林,王芳.温度和体重对中国对虾碳收支的影响[J].应用生态学报,2000,11(4):615-617.
[33] 李宝泉,杨红生,张涛,等.温度和体重对刺参呼吸和排泄的影响[J].海洋与湖泊,2002,33(2):182-185.
[34] 赵元凤,吕景才,刘婧.复合螯合微量元素添加剂对鱼、贝生长及能量转化的影响[J].吉林农业大学学报,2001,23(2):96-100.
[35] Cheng Zongjia J,Hardy R W,Usry J L.Plant protein ingredients with lysine supplementation reduce dietary protein level in rainbow trout(Oncorhynchus mykiss)diets and reduce ammonia nitrogen and soluble phosphorus excretion[J].Aquaculture,2003,218(1-4):553-565.
[36] 杨严鸥,谢綬启,熊邦喜,等.饲料质量对丰鲤和奥尼罗非鱼氮及能量收支的影响[J].水生生物学报,2004,28(4):337-341.
[37] 赵艳,童圣英,张硕,等.温度和盐度对虾夷马粪海胆耗氧率和排氨率的影响[J].中国水产科学,1998,5(4):33-36.
[38] 陈政强,单剑宇,陈昌生,等.九孔鲍耗氧率和窒息点的初步研究[J].水产科技情报,2001,28(1):11-14.
[39] 吴成业,阎希柱,谢綬启,等.不同规格黑脊倒刺鲃的耗氧率和排氨率初步研究[C]//中国水产学会第四届青年学术年会摘要集.北京:中国水产学会,2004:14-19.
[40] 温小波,库夭梅,罗静波.克氏原螯虾的生理生态学研究:I.标准代谢[J].湖北农学院学报,2003,23(1):41-44.
[41] 栗志民,刘志刚,谢丽,等.体重和温度对华贵栉孔扇贝(Chlamys nobilis)耗氧率和排氨率的影响[J].海洋与湖沼,2010,41(1):99-104.
[42] 黄东科,梁华芳.盐度和体质量对波纹龙虾耗氧率和排氨率的影响[J].广东海洋大学学报,2012,32(4):8-11.
[43] 李祖军,王道尊,龚希章.从能量代谢水平评价鱼用饲料的研究[J].上海水产大学学报,1997,6(3):186-192.
Effects of dietary selenium sources on respiratory metabolism of sea cucumber Apostichopus japonicus
Abstract:Juvenile and adult sea cucumber Apostichopus japonicus were fed the diets supplemented with seleniumethionine,sodium selenate and sodium selenite at a rate of 0.6 mg/kg for 60 days to study the effect of different selenium sources on sea cucumber specific growth rate and respiratory metabolism index.The results showed that all the sea cucumber fed the diets supplemented with selenium had significantly higher specific growth rate than the animals in the control group did,the maximal specific growth rate in the selenomethionine group,significantly higher than in the sodium selenate group and sodium selenite group(P<0.05).There was higher specific growth rate in the adults than that in the juveniles except for the selenomethionine group.All the sea cucumber fed the diets supplemented with selenium had significantly higher oxygen consumption rate than the animals in the control(P<0.05)did,the minimal oxygen consumption rate in selenomethionine group,significantlty lower than sodium selenate and sodium selenite groups(P<0.05),and the juvenilese higher than the adults.There was lower ammonia excretion rate in the sea cucumber fed the diets supplemented with selenium than in the control,the minimal in selenomethionine group,and significantly lower than in the control(P<0.05),without significant difference among sodium selenate group,sodium selenite group and the control(P>0.05).The juveniles had higher ammonia excretion rate than the adults did.There was significantly higher O/N value in the sea cucumber fed the diets supplemented with selenium than in the test group(P<0.05),the minimal O/N in selenomethionine group,without significant difference among all the treated groups(P>0.05),and he juveniles higher than the adults.The findings suggested that addition of selenium improved metabolic rate and growth,with the organic selenium(selenomethionine)being better than the inorganic selenium(sodium selenate,and sodium selenite)and juveniles being better than the adults in the selenomethionine group.
Key words:Apostichopus japonicus;selenium sources;specific growth rate;oxygen consumption rate;ammonia excretion rate;O/N
中图分类号:S963.16
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2016.01.011
文章编号:2095-1388(2016)01-0065-06
收稿日期:2015-05-19
基金项目:国家 “十二五”科技支撑计划项目 (2011BAD13B03)