中间球海胆(♀)与光棘球海胆(♂)种间杂交及自繁后代生长与表型特征比较
亓守冰,张伟杰,常亚青,田晓飞,王海峰,赵帅,经晨晨
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023)
摘要:为了解中间球海胆Strongylocentrotus intermedius(Si)与光棘球海胆Strongylocentrotus nudus(Sn)杂交后代的杂种优势及表型特征,以同期培育的中间球海胆(♀)与光棘球海胆(♂)种间杂交子一代(Si♀× Sn♂)幼胆及自繁子一代幼胆为研究对象,对3种海胆的成活率、生长表现 (生长速度和表型变异)和表型特征 (管足色素细胞数量、骨片形状、棘刺的颜色和长度)进行了比较。结果表明:经过90 d的养殖,中间球海胆和光棘球海胆的最终成活率均为100%,杂交海胆为97%,但3种海胆间无显著性差异 (P>0.05);在0~60 d的养殖范围内,杂交海胆具有最快的特定生长率 (4.00%/d),且显著高于中间球海胆(2.91%/d)和光棘球海胆 (3.15%/d)(P<0.05),而后两者之间无显著性差异 (P>0.05),60~90 d时,3种海胆的特定生长率之间均无显著性差异 (P>0.05);试验结束时,杂交海胆体质量的变异系数(76.12%)显著高于光棘球海胆 (63.81%)和中间球海胆 (52.05%)(P<0.05),表明种间杂交显著增加了后代的遗传变异;光棘球海胆管足中色素细胞数量最多,中间球海胆最少,杂交海胆的色素细胞数量介于父母本之间;杂交海胆棘刺为浅紫色,介于中间球海胆的白色和紫海胆的深紫色之间,而棘长较父母本细短;与棘色和管足颜色不同,杂交海胆骨片两端的突起产生了新的变异。研究表明,在幼胆期杂交海胆的表型特征明显区别于父母本,具有更快的生长速度和更高的变异水平,具有较高的育种价值。
关键词:中间球海胆;光棘球海胆;种间杂交;生长;变异;表型特征
中间球海胆 Strongylocentrotus intermedius又称虾夷马粪海胆,隶属于棘皮动物门、海胆纲、正形目、球海胆科,主要分布于日本北海道和俄罗斯远东地区的海域。1989年大连水产学院从日本将该种引入中国并人工繁育成功[1]。较之光棘球海胆Strongylocentrotus nudus、海刺猬Glyptocidaris crenularis等中国土著种类,该种海胆棘刺较短,易于进行筏式吊笼养殖,并且其生殖腺颜色、口感等品质远远优于光棘球海胆、海刺猬和马粪海胆Hemicentrotus pulcherrimus等中国海胆种类。目前,中间球海胆已成为中国的主要海胆养殖种类。
培育具备生长、抗逆和品质等性状优良的品种是提高养殖效益的重要手段。远缘杂交是水产动物中应用较为广泛的育种方法,其目的是能产生更大程度的遗传变异,使后代具备在生长和抗逆等经济性状方面的杂种优势。关于中间球海胆的种间杂交有较多报道,常亚青等[2]进行了中间球海胆与马粪海胆和光棘球海胆之间的杂交试验,得到了中间球海胆与马粪海胆的正反交后代,并观察到杂交后代的体表性状介于父母本之间。王丽梅等[3]得到了中间球海胆(♀)与光棘球海胆(♂)的杂交后代,经浮筏养殖,认为杂交后代棘长接近于母本,颜色多样,生长速度优于父母本。周遵春[4]研究认为,中间球海胆与光棘球海胆正反交后代的棘长介于父母本之间,颜色与父母本不同,浮筏养殖表明,中间球海胆(♀)与光棘球海胆(♂)的杂交后代生长速度较母本快。Ding等[5]获得了中间球海胆(♀)与光棘球海胆(♂)、紫海胆Anthocidaris crassispina (♂)的杂交后代,经过22个月的实验室养殖,中间球海胆(♀)与紫海胆(♂)杂交后代体质量的杂种优势率超过50%,而中间球海胆(♀)与光棘球海胆 (♂)杂交后代体质量的杂种优势率仅为0.32%。田晓飞等[6]通过研究不同温度下中间球海胆 (Si)和光棘球海胆 (Sn)杂交后代 (Sn♂× Si♀、Si♂×Sn♀) 和自繁后代 (Si♂×Si♀、Sn♂×Sn♀)的胚胎和幼体发育,认为杂交海胆Sn♂×Si♀在胚胎发育早期具有母本的热耐受性和生长劣势,但后期具备父本的生长优势。中间球海胆(♀)与光棘球海胆(♂)的种间杂交较易实现[1,5,7],且二者同属北方海域,亲本也较易获得,因此,这一种间杂交组合更可能应用于生产或育种。目前,对这一杂交组合的生长速度尚无完全一致的结论。 虽然王丽梅等[3]、 周遵春[4]、 Ding等[5]均认为这一杂交组合较母本具有生长优势,但对杂交后代与父本的生长比较却有截然相反的结论。并且,针对这一杂交后代的表型特征的报道也较为粗略,尚待进一步补充。
种间杂交是对中间球海胆进行遗传改良的重要途径,而培育具有生长和存活优势的杂交海胆则是海胆杂交育种中一个重要的阶段目标。杂交海胆相对于父母本的表型差异是鉴定后代是否为杂合子最简便易行的方法。为探明杂交海胆的生长特征,验证杂交海胆在生长上与双亲自繁后代相比较是否具有杂交优势,进一步了解杂交海胆相对于父母本的表型差异,本研究中,在实验室条件下对中间球海胆(♀)与光棘球海胆(♂)杂交后代及其父母本的成活率、生长表现 (生长速度和表型变异)和表型特征 (管足色素细胞数量、骨片形状和棘色)进行了全面的比较,以期为中间球海胆的远缘杂交育种和养殖研究提供借鉴。
1 材料与方法
1.1 材料
2013年8月在大连海洋大学农业部北方海水增养殖重点实验室育得中间球海胆(♀)与光棘球海胆(♂)杂交子一代、中间球海胆自繁后代、光棘球海胆自繁后代,并在相同实验室环境下养殖到4月龄。
1.2 方法
1.2.1 生长试验 取杂交海胆幼胆、中间球海胆幼胆和光棘球海胆幼胆各200枚,将每种幼胆随机等分为4组作为4个重复,每个重复内50枚海胆,放至140 L水槽 (77.50 cm×47.50 cm×38.40 cm)中,杂交海胆Si♀×Sn♂组分别标记为IN1、IN2、IN3、IN4;中间球海胆自繁后代组标记为 II1、 II2、II3、II4,光棘球海胆自繁后代组标记为NN1、NN2、NN3、NN4。将各组海胆在相同实验室条件下进行养殖,过量投喂石莼或海带,保持所有组海胆投喂同样的饵料。每天流水30 min,每3 d换水一次,测定流水换水前后的温度为 (11.30± 3.25)℃,盐度为31.62±0.30,溶解氧为 (6.20± 0.23)mg/L,pH为8.15±0.21。
1.2.2 表型特征观察 取壳径为 (12.0±0.5) mm的3种海胆各5枚,相同海胆放入同一个直径为10 cm加满水的培养皿中,等海胆伸出管足,用剪刀快速剪取赤道部的管足,用塑料滴管吸取管足放在载玻片上,用另一块载玻片压在表面,在显微镜下观察管足的色素细胞以及骨片特征并拍照。每个海胆取赤道部最长的3根棘刺,用镊子完整取下,放在表面光洁的白色板上,将3种海胆棘刺排列好,在解剖镜下观察并拍照。
1.2.3 指标的计算 试验开始时测量各重复组内所有海胆的初始体质量,之后每隔30 d测量所有海胆的体质量,试验持续90 d。试验海胆各重复组的特定生长率 (SGR)、变异系数 (CV)、存活率(SR)计算公式如下:
SGR(%/d)=100×(ln Wf-ln Wi)/t, CV(%)=100×(SD/W), SR(%)=100×(Nf/N)。
其中:Wf、Wi分别为各重复组每一阶段的终末和初始平均体质量 (g);t为该阶段时长 (d);SD为各重复组内体质量的标准偏差 (g);W为各重复组内体质量的平均值 (g);Nf为组内成活个体数 (尾);N为组内个体总数 (尾)。
1.3 数据处理
用Excel 2007软件对试验数据进行初步整理。利用SPSS 13.0软件,以初始体质量为协变量,对不同种类海胆在各养殖阶段的体质量及其特定生长率和变异系数进行协方差分析,分析前保证数据的正态性。种类影响达显著水平时进行Duncan多重比较。显著性水平设为0.05。
2 结果与分析
2.1 生长性状
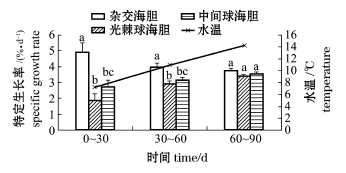
表1为3种海胆从试验开始至结束时各养殖阶段的体质量、变异系数和存活率。从表1可见:试验开始时,杂交海胆的体质量显著高于中间球海胆和光棘球海胆自繁后代 (P<0.05);在初始体质量作为协变量的情况下,试验至30 d时,杂交海胆的体质量显著高于父本光棘球海胆 (P<0.05),但与母本中间球海胆自繁后代无显著性差异 (P>0.05);试验至60 d和90 d时,3种海胆的体质量间均无显著性差异 (P>0.05)。光棘球海胆和中间球海胆的变异系数随着试验的进行呈降低的趋势, 90 d时杂交海胆最终的变异系数较初始时高,且显著高于其父本和母本 (P<0.05)。3种海胆的存活率自始至终无显著性差异 (P>0.05)。
表1 3种海胆在各试验阶段的体质量、变异系数和终末成活率 (mean±S.D.,n=4)
Tab.1 Body weight,coefficient of variation and the final survival rate of sea urchin Strongylocentrotus nudus,S.intermedius and the hybrids in each stage of the experiment(mean±S.D.,n=4)
注:同列中标有不同小写字母者表示组间有显著性差异 (P<0.05),标有相同小写字母者表示组间无显著性差异 (P>0.05)
Note:Themeans with different letterswithin the same column are significantdifferences at the0.05 probability level,and themeanswith the same letters within the same column are not significant differences
种类species 0 d 30 d 60 d 90 d体质量BW/g变异系数CV/%体质量BW/g变异系数CV/%存活率SR/%体质量BW/g变异系数CV/%存活率SR/%体质量BW/g变异系数CV/%存活率SR/%杂交海胆Si♀×Sn♂0.11± 0.05a65.91± 11.35a0.29± 0.04a78.00± 2.60a 100a 0.65± 0.07a79.24± 8.26a 100a 1.70± 0.11a76.12± 3.59a 97a光棘球海胆自繁S.nudus 0.06± 0.01b77.26± 11.35a0.15± 0.03b80.70± 2.60a 100a 0.48± 0.05a79.50± 8.26a 100a 1.78± 0.08a63.81± 3.59b 100a中间球海胆自繁S.intermedius 0.06± 0.02b78.39± 11.35a0.18± 0.02ab78.40± 2.60a 100a 0.52± 0.04a65.56± 8.26a 100a 1.86± 0.07a52.05± 3.59c 100a
剔除协变量 (初始体质量)的影响后,3种海胆在各试验阶段的特定生长率估计值见图1。从图1可见:在0~60 d范围内,杂交海胆具有最快的特定生长率 (4.00%/d),显著高于中间球海胆(2.91%/d)和光棘球海胆 (3.15%/d) (P<0.05),后两者之间无显著性差异 (P>0.05); 60~90 d时,3种海胆的特定生长率均无显著性差异 (P>0.05)。
2.2 表型特征
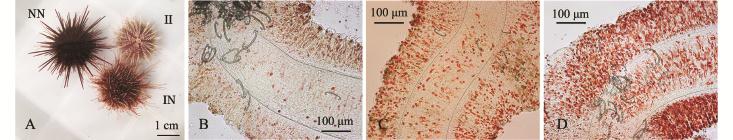
2.2.1 管足色素细胞数量和骨片 图2-A~D分别为中间球海胆、杂交海胆和光棘球海胆及其管足压片照片。从图2可见:3种海胆的色素细胞多分布于两侧,中间相对较少;中间球海胆管足中色素细胞数量最少 (图2-B),杂交海胆的色素细胞数量 (图2-C)介于父母本之间,这使杂交海胆的管足颜色介于两亲本之间;光棘球海胆的色素细胞数量最多 (图2-D)。
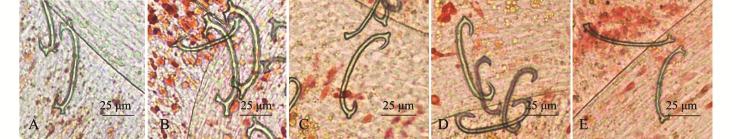
3种海胆管足中的骨片大都分布于管足末端,形状大致如拉长的字母 “C”,3种海胆管足骨片形状大致相同,仅在两端有细微差异。中间球海胆在骨片上、下两端各有两个短小的突起,且两个突起的间隔较小 (图3-A);光棘球海胆骨片上、下两端也各存在两个突起,但两个突起大小差异较大,且间隔较远 (图3-B);杂交海胆骨片上、下两端的突起都只有一个,并且突起较小,有一些骨片的突起消失,还有一些骨片退化成棒状 (图3-C~E)。
2.2.2 棘刺颜色与长度形态 图4为解剖镜下观察到的3种海胆的棘刺颜色和长度。从图4可见:中间球海胆棘色为白色,光棘球海胆为深紫色,而杂交海胆为浅紫色,介于中间球海胆和紫海胆之间;杂交海胆的棘长度与母本中间球海胆相近,但明显较细,光棘球海胆的棘刺明显较前二者长且较粗。
3 讨论
3.1 生长表现
试验动物的初始体质量往往会对试验期间的生长造成影响,表现为初始体质量作为协变量时对因变量具有显著影响[8-10]。协方差分析能在剔除协变量影响的情况下正确评估试验因素的影响。本试验中,3种海胆的初始体质量有显著性差异 (表1),因此,对各试验阶段体质量、特定生长率和变异系数等进行分析时,将其作为协变量,以避免得到不可靠的试验结论。另外,初始体质量不等,养殖密度大等因素会带来种内竞争,进而形成的环境内社会等级也是影响动物生长的重要因素[11-13],这使得从同一养殖空间 (如同一网箱)内取样作为重复的独立性不成立,从而影响数据分析的准确性。本研究中,将每种海胆养殖于4个水槽,水槽间海胆不形成社会等级关系,因此,利用水槽内海胆体质量平均值作为重复进行方差分析,符合独立性原则。
在养殖0~60 d内,杂交海胆较其父母本有较大的体质量和较快的生长速度 (SGR),表明在此阶段内,杂交后代具有超亲优势。这与王丽梅等[3]在浮筏养殖的试验结论一致。Ding等[5]获得的中间球海胆(♀)与紫海胆(♂)杂交后代也具有超亲优势;Wang等[14]报道了海湾扇贝和紫扇贝的种间杂交能产生生长速度优于双亲的杂交后代;张跃环等[15]研究认为,太平洋牡蛎(♀)与近江牡蛎(♂)杂交后代在幼虫期和稚贝期间也具有超亲优势。本研究与上述研究不同的是,杂交海胆在60~90 d养殖阶段内生长速度变慢,尽管3种海胆在这一阶段的体质量特定生长率与第90天时的体质量间均无显著性差异 (P>0.05),但杂交海胆的终末体质量已略低于其父母本。出现这一现象可能是因为杂交海胆的表型变异远大于中间球海胆和光棘球海胆 (表1),群体内竞争更为激烈,从而对小群体的平均生长速度产生了更大的影响[12-13]。另外,这一阶段的水温升高 (11.0~14.3℃)也可能对3种海胆的生长速度有不同程度的影响。陈林等[16]报道了杂交罗非鱼 (吉奥罗非鱼)具有显著高于父本的存活率,王丽梅等[3]试验发现,Si♀× Sn♂养殖成活率比其母本高10%,而本研究中3种海胆的存活率不存在显著性差异 (P>0.05),这可能是由本试验持续时间短、海胆个体较小且未度过夏季高温期等多重因素造成的。
增加群体的遗传变异是杂交的遗传学效应之一。孙振兴等[17]研究认为,盘鲍与皱纹盘鲍的杂交后代较皱纹盘鲍具有更高的变异系数。陈林等[16]研究发现,杂交罗非鱼的体质量变异系数高于双亲。以往对海胆种间杂交的研究中,对生长变异的关注较少,仅见常亚青等[2]报道了杂交海胆的壳径差异较大。本研究中首次对杂交海胆及其双亲对照的表型变异进行了分析。在一致的养殖环境下,杂交海胆的体质量变异系数显著高于两个亲本自繁后代,这表明杂交显著增加了后代的遗传变异水平。变异是育种工作的基础,在丰富的遗传变异中更容易选择出有利变异,从而培育出具备优良生产性状的品种,因此,杂交海胆具备了较高的育种价值。
3.2 表型特征
王丽梅等[3]、周遵春[4]研究发现,中间球海胆与光棘球海胆杂交后代的棘刺特征介于两亲本之间,且与母本相近,即母本体表为土黄色或黄褐色,棘短且较密;父本壳体表及大棘为黑紫色,棘长且较稀疏;杂交子一代体表为粉红色,棘较短且稍密。Ding等[5]认为,杂交海胆与所有亲本并无完全相同的形态特征,而是融合了父母本形态的结合体。本研究结果与上述研究结果基本一致,杂交海胆的棘色与亲本自繁后代存在显著性差异,中间球海胆棘色为白色,光棘球海胆为紫黑色,而杂交海胆为浅紫色,介于中间球海胆和光棘球海胆之间。稍有不同的是,本研究中杂交海胆的棘刺较中间球海胆更细短,中间球海胆棘色为白色而不是土黄色和黄褐色,其原因可能是本试验中所用的海胆处于幼胆阶段,不同于王丽梅和周遵春等所用的成胆,随着海胆的生长,其棘色和棘刺长度还有所变化。另外,本研究中首次对杂交海胆的管足颜色进行了显微观察,发现杂交海胆管足中的色素细胞数量介于两个亲本之间,与棘刺颜色类似,是父母本的中间类型。与管足和棘刺不同的是,杂交海胆的骨片形状尤其是骨片两端的突起并非父母本的中间体。中间球海胆和光棘球海胆的骨片上、下两端均各有两个突起,而杂交海胆的上、下两端的突起都只有一个,有一些骨片的突起甚至完全消失,这说明杂交产生了新的变异,这一变异的产生原因和研究价值有待进一步探讨。
参考文献:
[1] 常亚青,丁君,宋坚,等.海参、海胆的生物学研究与养殖[M].北京:海洋出版社,2004.
[2] 常亚青,王子臣,宋坚,等.四种海胆杂交的可行性及子代的早期发育[J].水产学报,2000,24(3):211-216.
[3] 王丽梅,韩家波,董颖,等.中间球海胆与光棘球海胆杂交子一代的生长比较研究[J].水产科学,2004,23(2):1-3.
[4] 周遵春.中间球海胆与光棘球海胆的杂交及分子遗传学的研究[D].青岛:中国海洋大学,2006.
[5] Ding J,Chang Y,Wang C,et al.Evaluation of the growth and heterosis of hybrids among three commercially important sea urchins in China:Strongylocentrotus nudus,S.intermedius and Anthocidaris crassispina[J].Aquaculture,2007,272(1):273-280.
[6] 田晓飞,常亚青,张伟杰,等.中间球海胆、光棘球海胆自繁和杂交后代幼体发育及对高温的反应[J].大连海洋大学学报, 2014,29(5):431-438.
[7] 王丽梅,韩家波,许伟定,等.中间球海胆与光棘球海胆杂交及子一代人工育苗技术[J].水产科学,2003,22(2):9-11.
[8] Jobling M,Wansvik A.Effect of social interaction on growth rates and food conversion efficiency of Arctic charr,Salvelinusalpinus L. [J].Journal of Fish Biology,1983,19:245-257.
[9] 姚峰,甄恕其,何爱华,等.初始体重差异对黄颡鱼补偿生长的影响[J].淡水渔业,2008,38(5):65-68.
[10] 崔正贺,王岩.不同体重异育银鲫(Carassiusauratusgibelio)的补偿生长[J].海洋与湖沼,2007,38(1):8-14.
[11] Brett JR.Environmental factors and growth[J].Fish Physiology, 1979(8):599-675.
[12] Jobling M,Baardvik BM.The influence of environmentalmanipulations on inter-and intra-individual variation in food acquisition and growth performance of Arctic charr,Salvelinus alpinus [J].Journal of Fish Biology,1994,44(6):1069-1087.
[13] Irwin S,O'halloran J,Fitzgerald R D.Stocking density,growth and growth variation in juvenile turbot,Scophthalmus maximus (Rafinesque)[J].Aquaculture,1999,178(1/2):77-88.
[14] Wang C,Liu B,Li J,et al.Introduction of the Peruvian scallop and its hybridization with the bay scallop in China[J].Aquacul-ture,2011,310(3/4):380-387.
[15] 张跃环,王昭萍,闫喜武,等.太平洋牡蛎与近江牡蛎的种间杂交[J].水产学报,2012,36(8):1215-1224.
[16] 陈林,李思发,简伟业,等.吉奥罗非鱼(新吉富罗非鱼♀×奥利亚罗非鱼♂)生长性能的评估[J].上海水产大学学报, 2008,17(3):258-262.
[17] 孙振兴,常林瑞,宋志乐.皱纹盘鲍与盘鲍杂交效果分析[J].水产科学,2005,24(8):1-3.
Grow th and phenotypic characteristics of interspecific hybrids of sea urchin Strongylocentrotus intermedius(♀)× S.nudus(♂)and purebred offsprings
QIShou-bing,ZHANGWei-jie,CHANG Ya-qing,TIAN Xiao-fei, WANG Hai-feng,ZHAO Shuai,JING Chen-chen
(Key Laboratory of Mariculture&Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China)
Abstract:The survival,growth performance[specific growth rate(SGR)and coefficient of variation(CV)]and phenotypic characteristics(count of chromatophore cells and ossicle shape on tube feet,and spine length and color)were compared in sea urchin hybrid juveniles of Strongylocentrotus intermedius(♀)×S.nudus(♂)and the two purebred sea urchin offsprings to evaluate the heterosis.The 90 d feeding trial showed that both S.intermedius and S.nudus had survival rate of 100%,and the hybrids 97%at the end of the experiment,without significant difference(P>0.05).However,there was significantly higher SGR(4.00%/d)in the hybrids than that in the parents(2.91%/d in S.intermedius and 3.15%/d in S.nudus)(P<0.05),without significant difference between the two parental species(P>0.05).From 60 days to 90 days,therewas no significant difference among the three sea urchins(P>0.05).The hybrids had significantly higher coefficient of variation for body weight of (76.12%)than the S.intermedius(52.05%)and S.nudus(63.81%)at the end of the experiment(P<0.05), indicating that interspecific hybridization significantly led to increase in genetic variation in hybrid offsprings.The maximal chromatophore cellswere observed on the tube feet in S.nudus while theminimal oneswere found in S.intermedius.The hybrids showed themoderate number of chromatophore cells on tube feet.The hybrids had spines with a light purple color,between S.intermedius(white)and S.nudus(deep purple),and thinner and shorter spines compared to the parental sea urchins.Unlike the colors of spines and tube feet,a new variation was observed at the ends of the ossicles on tube feet.The findings indicate that the hybrid juveniles has an obviously different phenotype from that of the parents,and are characterized by good growth rate and higher level of variation, as a high breeding potential.
Key words:Strongylocentrotus intermedius;S.nudus;hybrid;growth;variation;phenotypic characteristics
中图分类号:S917.4
文献标志码:A
收稿日期:2014-11-23
基金项目:辽宁省农业攻关及成果产业化项目 (2014203005);国家 “863”计划现代农业技术领域重大项目 (2012AA10A412)
作者简介:亓守冰 (1989—),硕士研究生。E-mail:qishoubing521@126.com
通信作者:常亚青 (1967—),博士,教授。E-mail:yqchang@dlou.edu.cn
DOI:10.16535/j.cnki.dlhyxb.2015.05.002
文章编号:2095-1388(2015)05-0461-06