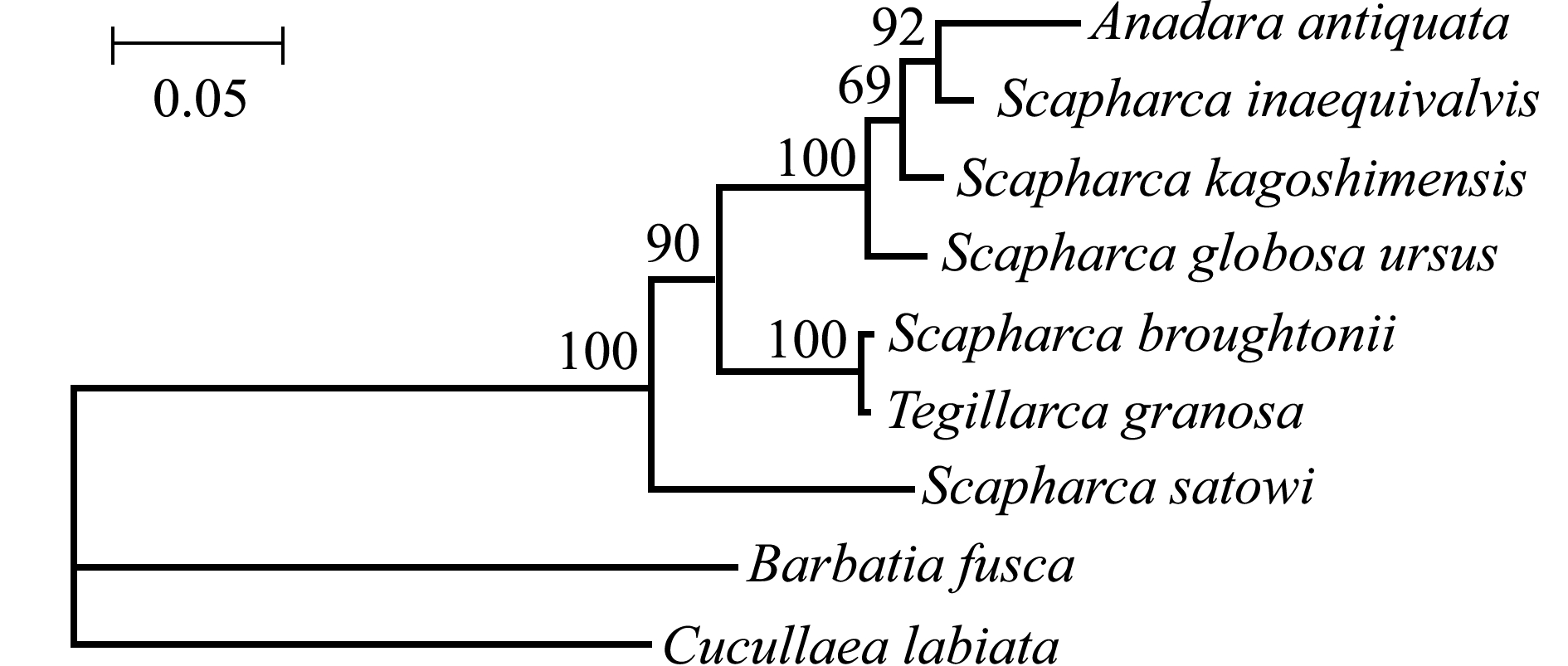
图1 蚶科7种贝类的系统进化与亲缘关系树
Fig.1 Neighbor-joining tree of 7 species in Arcidae inferred from them tDNA COI genes
摘要:毛蚶Scapharca subcrenata是一种重要的海产经济贝类,受过度捕捞及生境破坏等因素影响,近年来其资源和产量大幅度下降。本研究中综述了毛蚶的生物学特性、人工繁育、生理生态和遗传学等相关研究概况,阐明了对毛蚶的栖息环境、繁殖周期、胚胎发育、人工养殖、生理生态、系统进化、种群分化等方面的研究现状,并对其发展前景进行了展望。本研究可为开展毛蚶增殖放流、资源修复、人工养殖、遗传多样性保护等研究与政策制定提供科学依据。
关键词:毛蚶;生物学特性;人工繁育;人工增养殖;遗传学
毛蚶Scapharca subcrenata隶属于软体动物门Mollusca、双壳纲Bivalvia、翼形亚纲Pteriomorphia、蚶目Arcoida、蚶科Arcidae、毛蚶属Scapharca,俗称毛蛤、麻蛤、麻蚶、瓦楞子等[1],是重要的海产经济食用贝类,其不仅肉味鲜美,具有高蛋白、低脂肪、高维生素含量等特点,还具有药用价值,深受消费者喜爱。20世纪60—80年代,毛蚶资源量和采捕数量都很大,仅渤海湾资源蕴藏量就曾达数十万吨,1976年河北省毛蚶产量达到最高水平4.1万t,不仅是渔业生产的重要捕捞对象,也是重要的出口创汇品种[2-3]。进入20世纪80年代,受过度捕捞及生境破坏等因素影响,毛蚶资源和产量开始大幅度下降,已远远不能满足市场需求。近年来,毛蚶的资源恢复和增养殖越来越受到重视,对毛蚶的基础性研究和苗种繁育、养殖生产等工作得到了有效开展[4-7]。本研究中,综述了毛蚶的生物学、人工繁育、增养殖和遗传学等相关研究进展,以期为进一步开展毛蚶的生物学研究提供参考,并推动其资源修复与增养殖工作的深入开展。
1.1 资源分布与栖息环境
毛蚶广泛分布于东亚的中国、日本、朝鲜沿岸[8-9],在中国以莱州湾、渤海湾、辽东湾、海州湾等浅海区资源尤为丰富,辽宁锦州、河北唐山、天津北塘、浙江象山港和山东羊角沟等均是毛蚶的重要产地[1]。毛蚶主要栖息于浅海 (20m以下)的软泥底或含砂的泥质海底,栖息水域一般是稍有淡水流入的内湾和较平静的浅海,多分布在水深5~7m的海域,水深14m以上的海域分布稀少,由沿岸浅水向外海深水呈密度递减趋势[10]。潮间带和潮下带也有一定数量的分布,潮间带的毛蚶整个卧于泥沙中,仅留一出入水口;而潮下带的毛蚶仅三分之一躯体卧于泥沙中。毛蚶主要以圆筛藻、舟形藻等一些硅藻类浮游植物、桡足类附肢、双壳类卵、浮游幼体和有机碎屑等为食,对温度和盐度的适应范围较广,适宜生存盐度为 21.0~28.8,存活水温为0~32℃[5-6]。
1.2 形态特征
毛蚶贝壳中等大小,壳质坚厚,双壳膨凸,背侧两端略显棱角,腹缘前端圆,后端稍延长,通常两壳大小不等,右壳稍小,壳顶突出并向内卷曲,位置偏向前方。壳面放射肋突出且较密,有30~34条放射肋,肋上有方形小结节,状似瓦垄,在左壳尤为明显,腹侧有明显的生长纹。外壳覆盖着一层深褐色绒毛,壳面和壳内均为白色。铰合部呈直线形,齿密而小,约50枚[5-7]。
形态学分析是物种分类学研究的常用方法,也是生物起源与进化研究、物种分类、资源鉴定的基础。 陈丽梅等[11]、 郝伟等[12]、 王辉等[13]用通径分析法研究了形态性状对毛蚶体质量的影响,得出壳长和壳高是影响体质量的主要因素。李明云等[14]研究发现,渤海湾毛蚶与象山港毛蚶在形态学和生化组成上存在明显差异。陈蓉等[15]用多变量形态度量学分析方法,比较了中国5个海区的5个野生群体毛蚶的形态差异,发现天津塘沽和山东青岛群体形态最为接近,广西北海群体趋异程度最大。宋菲菲等[16]用13个形态性状对山东省4个地区及辽宁省葫芦岛和海南省三亚6个群体间的形态差异进行了比较,发现东营河口和东营广饶群体形态最为接近,海南群体趋异程度最大。因此,形态差异比较可以为毛蚶的种群鉴别与种质资源保护,以及研究群体间形态变异与环境间的关系提供基础数据。
1.3 繁殖生物学
毛蚶大多为雌雄异体,个别为雌雄同体,生殖腺包在消化腺 (内脏团)周围,性成熟时雌性性腺为桔红色,雄性性腺为乳白色,生殖孔开口在后闭壳肌的腹面,左右各1个。阎斌伦等[17]、Yurimoto等[18]、Park等[19]根据组织学和外部形态特征分别研究了毛蚶在中国连云港、日本Ariake湾和韩国Yeoja湾的性腺发育和繁殖周期,将生殖腺发育主要分为5个时期 (增殖期、生长期、成熟期、排放期、休止期)或4个时期 (生长期、成熟期、排放期、排尽期)。连云港毛蚶性腺发育的5个时期分别为:增殖期 (3—4月),生殖腺发育不明显,肉眼不能分辨雌雄;生长期 (5—6月),性腺发育明显,肉眼可辨雌雄;成熟期 (6—7月),性腺几乎充满整个内脏团并延伸到足的基部;排放期(7—9月),排放初期有少量排放,肉眼无法判断是否已排放,大量排放后的性腺饱满度明显下降,肉眼很容易鉴别;休止期 (10月—翌年2月),软体部消瘦,性腺不明显,不能分辨雌雄[17,20]。
毛蚶繁殖方式为卵生型,产卵量较大,壳长为3~4cm的雌蚶一次可产卵250~300万粒。繁殖期间,雌、雄个体分别将成熟精卵从生殖孔排出体外,在海水中完成受精,浮游一段时间后附着变态为稚贝,再进一步发育为成体。受水温、饵料等环境因素影响,不同海区的毛蚶繁殖期略有差异,但大都集中在7—9月,当水温达到22~24℃时开始产卵,水温为26.7℃左右进入繁殖盛期[17-19]。辽东湾毛蚶的繁殖季节为7月上旬至8月末,此时水温为22~27℃,产卵盛期在7月15日—30日,水温为24~26℃[21]。毛蚶生殖腺在繁殖期具有分批成熟、分批排放的特点,第一次排放量最大,高峰间隔约半个月,一般可排放3~4次[22]。
竺俊全等[23]对毛蚶精子超微结构观察发现,其精子顶体为圆锥形,精核无核前窝,具核后窝;中段横切面常见 4~5个线粒体环绕于中心粒周围,轴丝为典型的 “9+2”结构。精子发生过程中线粒体的行为可分为精原细胞期、初级精母细胞期、次级精母细胞期和精母细胞期[24]。董迎辉等[25]对毛蚶受精和卵裂过程中核行为的细胞学变化观察发现:在水温为 (26±1)℃时,受精卵在15~18、20~25min先后排出第一、第二极体,完成第一、第二次成熟分裂;受精约30min时,雌、雄原核形成;35min时,雌、雄原核各自形成染色体组;40min时,受精卵第一次卵裂成2个大小不等的卵裂球;55~60min时,受精卵第二次卵裂成1大3小4个卵裂球。朱东丽等[26]对毛蚶 (♂)×泥蚶 (♀)杂交受精细胞学观察发现:毛蚶精子可以穿过泥蚶卵膜进行受精,使其释放第一极体和第二极体;受精卵可以进行卵裂,发育速度较自交组慢;杂交胚胎前两次卵裂正常,后期出现畸形,囊胚仅能发育至担轮幼虫期。
2.1 人工繁育
近几年,各地广泛开展了毛蚶人工繁育相关研究, 辽宁[27-31]、 河北[32-34]、 山东[35-36]、 江苏[37]和浙江[38-41]等地不同海区的毛蚶人工繁育和中间培育技术均有较多相关报道,人工繁育技术也越来越成熟。根据生产需要可选择人工升温育苗或常温育苗,一般包括亲蚶采捕、亲蚶培育、催产孵化、幼虫培育、附着基投放和稚贝培育等步骤。翟林香等[4]、马云聪等[5]、 徐国成等[37]对毛蚶的人工育苗技术进行了总结,包括亲蚶培育、幼虫培育和稚贝培育等关键技术环节。
沈伟良等[41]研究了温度与盐度对毛蚶受精卵孵化及幼虫生长的影响,受精卵孵化的适宜温度为25~30℃,适宜盐度为18~30,幼体生长的适宜温度为25~30℃,适宜盐度为20~25。通过研究饵料对毛蚶浮游幼虫生长、发育、存活和体内脂肪酸组成变化的影响发现,不同饵料对幼虫生长、存活和附着变态的影响存在差异:单独投喂时,金藻Isochrysissp.效果较好,小球藻Chlorellasp.效果最差;混合投喂时,金藻与其他饵料混合投喂效果较好[31,39-40,42]。 一些研究还比较了聚乙烯网片、 棕帘、波纹板和扇贝壳等附着基对幼虫附着变态的影响,发现聚乙烯网片附着效果最好,附着基的放置方式和水层对附着效果也有显著影响[34,36,43-44]。
人工繁育的毛蚶苗种,由于规格较小,成活率不够稳定,需要进行适当的中间培育,使苗种达到一定规格之后再进行放养才具有较高的成活率。黄金臣等[45]利用室内水泥池进行了稚贝中间培育试验,当壳长达到0.5cm之后在虾池进行后期培育,取得了较好效果。葛立军等[46]研究了毛蚶稚贝池塘中间培育的适宜密度,发现中间暂养首先以60目网袋为宜,并应在不同阶段适时分苗。王海涛等[47]通过升温育苗使稚贝出池时间比常温育苗提前1~2个月,延长了稚贝的生长时间,有效提高了苗种的越冬成活率。孙同秋等[48]利用虾池围网进行了毛蚶苗种的高密度中间培育,培育成的养殖用大规格苗种,有利于翌年集中捕捞、提早放苗。
2.2 半人工采苗
半人工采苗是根据毛蚶幼虫在附着变态阶段具有用足丝附着生活的习性,在繁殖季节向毛蚶渔场投放适宜的附着基采集天然苗种。开展半人工采苗的海区一般应具备亲蚶充足、潮流畅通、流速适宜、底质与水质适宜、饵料丰富、敌害生物少等条件[5]。20世纪80年代已有在渤海湾开展毛蚶半人工采苗的相关报道,研究者通过测定性腺指数和监测幼虫数量与个体发育两种方法进行采苗预报,在适宜时机投放附着基。结果表明,表层和底层采苗的数量差异不明显,各种附着基都能采到一定数量的稚贝,其中聚乙烯丝团或网片的数量最多,这可能与聚乙烯网表面粗糙、间隙多,稚贝附着后不易被水流带走有关[22,49-50]。
2.3 池塘养殖
毛蚶的池塘养殖一般采用池塘稚贝挂袋或笼吊暂养、室内控温越冬、池塘底质处理和水质调控等方式进行。利用毛蚶与对虾、鱼类栖息水层、食性等生态学与生物学特性差异开展虾贝立体养殖,形成互利共生的池塘生态环境,防止池塘富营养化,提高池塘养殖的综合经济效益[51]。目前,毛蚶与对虾的配套混养技术较为成熟,对虾品种可根据实际情况灵活选择。刘祖祥等[52]和刘洪文[53]开展的对虾与毛蚶池塘混养的试验表明:毛蚶的滤食性摄食可消耗虾塘中大量的浮游植物和对虾残饵,改善水质环境,促进了对虾的生长;同时由于虾塘的环境较稳定(不受潮汐涨落的影响)和饵料充足等原因,毛蚶的生长也较快。孙同秋等[54]在黄河三角洲沿海池塘开展了毛蚶与日本对虾Penaeus japoni+ cus、脊尾白虾Exopalaemon carinicaud和梭鱼Che+ lon haematocheilus的立体养殖技术研究,实现了减轻养殖污染排放、提高经济与环境效益的目的。
2.4 浅海底播与筏式养殖
浅海滩涂底播增殖和浮筏养殖双壳贝类在中国有着悠久的历史,毛蚶、魁蚶、泥蚶等蚶科贝类都是常见的养殖品种。付卓等[55]通过对锦州浅海毛蚶底播区底质环境与增殖效果调查,证实了在锦州浅海开展毛蚶种群资源修复与高效增殖的可行性。张建春等[50]利用扇贝养殖器材摸索出一套成功的毛蚶半人工采苗及筏式笼养技术;战江祥等[56]在海州湾进行了毛蚶筏式养殖试验,获得了一定的成功经验;林培振等[57]在大连金州海区连续两年开展毛蚶筏式笼养生产性研究,取得了较好的经济效益。此外,日本也广泛开展了毛蚶筏式吊养与底播笼养试验,相关研究均表明,毛蚶在无底质环境条件下进行浮筏式笼养或沉入式吊养是可行的,但在养殖过程中需对养殖笼(筐)的深度进行适时调整。研究还发现,毛蚶的生长和存活,在夏季主要受溶解氧水平的影响,在冬季主要受水温的影响[58-60]。
3.1 温度、盐度的适宜性
毛蚶对温度的适应性非常广,在水温为2~30℃的条件下均能生存,其生长的适宜温度为18~28℃。与其他变温动物一样,温度的高低对毛蚶新陈代谢具有十分重要的影响。姜祖辉等[61]研究了温度、规格对毛蚶耗氧率和排氨率的影响,发现在一定温度范围内,毛蚶的耗氧率和排氨率与温度呈正相关关系,与壳长呈负相关关系。毛蚶能根据海水盐度的变化,通过渗透压调节满足其正常的生理活动。沈伟良等[62]对不同盐度下毛蚶稚贝耗氧率和排氨率的研究表明:耗氧率和排氨率随盐度的升高而升高,当盐度达到30后开始下降;O∶N值也随着盐度的升高而升高,在盐度达到30后开始下降。史宝等[63]对不同盐度下不同规格毛蚶成贝耗氧率和排氨率的研究表明,单位体质量的耗氧率和排氨率在盐度32时达到最大,而后随着盐度的升高而逐渐降低。施祥元等[64]报道了毛蚶稚贝的适宜盐度范围为17~37,最适盐度范围为27~33。鈴木秀幸等[65]报道了在自然海区低盐度 (7~9)条件下,毛蚶的存活天数不足15d。
3.2 溶解氧、pH、氨氮的适宜性
溶解氧、pH、氨氮也是影响水生生物生存的重要环境因子。本田匡人等[66]研究了低溶解氧(<0.5mg/L)对毛蚶呼吸代谢的影响,结果表明,在低溶解氧环境下,毛蚶软体部的苹果酸、丙酮酸和反丁烯二酸含量下降,丙酸含量显著提高;低溶解氧对毛蚶呼吸代谢的促进作用,可导致其存活率下降。鈴木秀幸等[65]报道,在夏季自然海区低溶解氧 (<2mg/L)环境下,毛蚶的存活时间约为2周。方军等[67]研究了 pH和氨氮对毛蚶稚贝生长和存活的影响,结果表明,稚贝适宜生存的pH范围为7.5~8.5,pH为8.0时生长与存活最好;稚贝虽然对氨氮具有较高的耐受性,但氨氮浓度越高存活率越低,对氨氮24、48、72、96h的半致死浓度分别为96.33、74.16、43.63、20.36mg/L。
3.3 摄食与滤水
由于具有滤食能力和生物沉积的特性,双壳贝类在维持海洋物质循环和生态平衡方面发挥着重要作用。Nakamura[9]研究了日本Ariake湾毛蚶的滤水率 (CR),结果表明,毛蚶的CR与软体部干质量呈指数性相关;在10~20℃下,CR随温度的升高而升高;饵料浓度 (叶绿素含量为 10~40 mg/L)和日供饵料量 (干质量的 0.5%/d~6 %/d)对CR均无显著影响。沈和定等[68]研究了不同水温下毛蚶对5种单细胞藻类的滤除率、选择性和表观消化状况,结果表明,毛蚶不仅对不同大小、形状的藻类具有选择性,而且对大小、形状相似的藻类也具有明显的选择性;水温22℃下,毛蚶对5种藻类的表观消化状况不同。杨凤等[69]进行了温度、盐度和充气对毛蚶排遗及泥沙净化影响的试验,结果表明,在盐度为30的条件下,不同温度组的毛蚶均能正常排遗,10h内排遗速率以20℃时最高;在20℃条件下,不同盐度组毛蚶均能正常排遗,10h内排遗速率以盐度为25时最大;不充气条件下大于充气条件下的排遗速率。
4.1 染色体组型
自20世纪80年代开始,有关毛蚶的染色体特征、系统进化、种群分化和分子标记开发等遗传学特性相关研究已陆续开展。和田克彦等 报道了毛蚶的染色体组型,毛蚶的二倍体染色体数目为2N=38。陆荣茂等[71]以担轮幼虫为材料,比较了毛蚶、泥蚶 Tegillarca granosa、橄榄蚶 Estellarca olivacea的核型差异,结果表明,3种蚶的染色体数目均为2N=38,但核型公式有所差异,分别为20m+12sm+6st、22m+12sm+4st和18m+18sm+2st,染色体臂数 (NF)分别为70、72、74,均未发现性染色体和随体。
4.2 系统进化
对毛蚶的系统进化研究多见于对毛蚶、泥蚶和魁蚶S.broughtonii的比较分析。孙桂金等[72]和张波等[73]分别利用ITS1和ITS2序列对3种蚶进行了系统发育分析研究,结果均显示,毛蚶和泥蚶的遗传关系较近。郭永军等[74]对3种蚶的核糖体 DNA转录间隔区 (ITS区)进行限制性片段长度多态性(RFLP)分析,发现6种限制性内切酶均能将泥蚶与毛蚶及魁蚶区别开来,但未发现毛蚶和魁蚶间特异性的RFLP标记。Tanaka等[75]利用4种线粒体基因 (COI、16S rRNA、12S rRNA和Cyt b),以棕蚶Barbatia fusca和粒帽蚶Cucullaea labiata为外类群,构建了毛蚶、泥蚶和魁蚶等7种蚶科贝类的进化关系树 (图1),为蚶科贝类的分类与进化研究在分子水平上提供了科学依据。
图1 蚶科7种贝类的系统进化与亲缘关系树
Fig.1 Neighbor-joining tree of 7 species in Arcidae inferred from them tDNA COI genes
4.3 遗传多样性与种群分化
研究毛蚶不同地理群体的遗传多样性及种群遗传分化状况,可以为毛蚶的种质资源保护、良种选育和合理开发等提供基础资料。李旭光等[76]利用同工酶技术对江苏海州湾、浙江象山港和辽宁辽东湾3个毛蚶群体的遗传多样性进行了研究,发现3个群体的遗传多样性仍较丰富,海州湾群体和象山港群体间遗传分化较小,辽东湾群体与其他两个群体间的遗传分化较大。王晓梅等[77]对塘沽、大连、宁波3个毛蚶群体的 rDNA转录间隔区进行了RFLP分析,表明3个毛蚶群体的遗传多样性水平较低,大连和塘沽样本的亲缘关系较近。赵文等[78]用ISSR技术研究了辽宁沿海毛蚶5个群体的遗传多样性,结果表明,5个群体的多态位点比例和平均杂合度都处于较高水平,但群体间的遗传分化水平较低。宋菲菲[79]利用核糖体ITS2序列和线粒体COI基因对山东沿岸毛蚶8个群体遗传多样性进行了分析,结果表明,不同群体间均存在单倍型类群,但单倍型类群在群体间的分布差异不显著,群体间遗传分化也不显著,遗传多样性和遗传分化水平较低可能与频繁的群体间基因交流有关。Tanaka等[80]利用线粒体COI基因研究了毛蚶8个日本群体和1个韩国群体225个体的遗传多样性和种群分化,共发现56个单倍型,其中包括14个共同单倍型,日本和韩国群体有3个相同的共同单倍型;单倍型和核苷酸多态性分别为0.65% ~0.93%和0.22%~59%,表明各群体处于较高的遗传多样性水平,但韩国群体的遗传多样性相对偏低。
4.4 分子标记开发
微卫星标记 (SSR)具有多态性高、共显性、DNA用量少、易检测、重复性好等优点,目前有关毛蚶分子标记开发的相关报道多集中在微卫星标记开发方面。Feng等[81]用磁珠富集法开发了14个毛蚶基因组微卫星,在40个野生个体中验证得到的平均等位基因数为11~29个,观测杂合度和期望杂合度分别为0.400~0.923和0.705~0.965。Li等[82]从魁蚶的表达序列标签 (EST)中开发了25个多态性较好的EST-SSR,并验证了这些标记在毛蚶和泥蚶中的通用性,结果表明,这些标记在毛蚶中的通用率达100%,在泥蚶中仅为20%。Dong等[83]从泥蚶中开发了62个多态EST-SSR,并验证了这些标记在毛蚶中的通用性,其中16个标记可在毛蚶中应用,通用率为25.81%。这些分子标记的开发为毛蚶的遗传多样性分析、种群分化、种质资源鉴定、遗传连锁图谱构建和数量性状定位等研究提供了技术保障。
5.1 开展种质资源保护与良种选育的研究
中国的毛蚶种质资源相对丰富,这些野生种质群体是人工繁育的宝贵基因库。但随着过度开发和生境破坏,毛蚶的野生资源日趋减少。从已有的报道来看,中国的毛蚶种质资源已呈日益衰退趋势。虽然近年来广泛开展的资源恢复和人工增养殖活动,使其种群数量有所回升,但也带来了一些问题:由于资源恢复和增养殖所用的苗种均为人工苗种,目前人工苗种的亲本都是野生毛蚶,养殖户为了调剂亲本不足和选择优质亲本,经常跨地区引进亲本,加之频繁的苗种交换,群体间遗传多样性和遗传分化水平日趋下降;同时很多育苗场不注重选育,有效繁殖群体过小,近交严重,导致毛蚶生长速度降低,抗逆性和抗病性越来越差。因此,应加强毛蚶种质资源保护和良种选育的基础和应用研究,筛选与生长抗病相关的功能基因,开发大量多态有效且与优势生产性状相关的分子标记,培育生长快、抗逆性强的优良品种。
5.2 开发多营养级复合的健康生态养殖技术
双壳贝类具有很强的滤水能力,能够过滤大量细小的颗粒物质,通过摄取浮游植物和有机颗粒,同化一部分有机质,以粪便或假粪的形式排出部分物质,在生态系统的物质循环和能量流动中发挥着重要作用。多营养级的综合养殖是近年提出的一种健康、可持续发展的海水养殖理念。对于资源稳定、守恒的系统,营养物质的再循环是生态系统中一个重要过程,由不同营养级生物,如滤食性贝类、投饵类动物、大型藻类和沉积食性动物等组成的综合养殖系统中,一些生物排泄到水体中的废物能够成为另一些生物的营养物质来源。因此,开展毛蚶等双壳贝类与对虾、鱼类和藻类的多营养级复合养殖研究,能充分利用输入到养殖系统中的营养物质和能量,把营养损耗及潜在的经济损耗降到最低,同时减小对环境的负面影响,包括减少污染物排放、改善养殖环境、减少有害菌、减少有害生物等,从而使系统具有较高的容纳量和经济产出。
参考文献:
[1] 王如才,王昭萍.海水贝类养殖学[M].青岛:中国海洋大学出版社,2008.
[2] 沈源远,顾亚卿,苗家禄.1979年渤海湾北部海区毛蚶资源调查报告[J].天津水产,1980(00):1-6.
[3] 王友智.渤海湾毛蚶资源现状及恢复资源措施的探讨[J].海洋渔业,1982(1):16-19.
[4] 翟林香,毛红茜.毛蚶的生物学特性及人工育苗技术[J].现代农业科技,2010(4):345-346.
[5] 马云聪,闫升华,苏文清,等.毛蚶育苗和养殖实用技术[M].北京:海洋出版社,2008.
[6] 陈建华,阎斌伦,高焕.毛蚶生物学特性及其研究进展[J].河北渔业,2006(9):24-25.
[7] 周军,张海鹏,李怡群,等.毛蚶生物学特性的研究[J].河北渔业,2005(2):18-24.
[8] Nakamura Y,Shinotsuka Y.Suspension feeding and growth of ark shell Anadara granos a:comparison with ubiquitous species Scapharca subcrenata[J].Fisheries Science,2007,73(4):889-896.
[9] Nakamura Y.Suspension feeding of the ark shell Scapharca subcre+ nata as a function of environmental and biological variables[J]. Fisheries Science,2005,71(4):875-883.
[10] 一九七六年毛蚶资源调查总结[J].水产科技情报,1977(3):1-7.
[11] 陈丽梅,刘利华,秦传新,等.渤海地区毛蚶形态性状对活体重的影响效果[J].安徽农业科学,2012,40(28):13813-13814.
[12] 郝伟,徐国成,庞作宽,等.毛蚶壳性状对体质量和软体部质量的影响效果分析[J].淮海工学院学报:自然科学版,2012, 21(3):81-87.
[13] 王辉,刘志刚,符世伟.南海毛蚶形态特征对体重的相关分析[J].热带海洋学报,2007,26(6):58-61.
[14] 李明云,冯建华,王尚友.渤海湾毛蚶与象山港毛蚶的种间差异[J].动物学杂志,1992,27(5):4-7.
[15] 陈蓉,刘建勇,唐连俊,等.中国5个海区不同群体毛蚶形态差异分析[J].海洋科学,2009,33(7):64-69.
[16] 宋菲菲,王其翔,刘名,等.不同地区毛蚶群体的形态差异的比较研究[J].中国海洋大学学报:自然科学版,2012,42 (11):40-45.
[17] 阎斌伦,许星鸿,郑家声,等.毛蚶的性腺发育和生殖周期[J].海洋湖沼通报,2005(4):92-98.
[18] Yurimoto T,Mori Y,Ito S,etal.Reproductive cycle of the subcrenated ark shell Scapharca kagoshimens is(Tokunaga,1906)in Ariake Bay,Japan[J].Journal of Shellfish Research,2008,27 (5):1101-1108.
[19] Park H J,LeeW C,Choy E J,et al.Reproductive cycle and gross biochemical composition of the ark shell Scapharca subcrenata (Lischke,1869)reared on subtidalmudflats in a temperate bay of Korea[J].Aquaculture,2011(322/323):149-157.
[20] 许星鸿,阎斌伦,徐国成,等.毛蚶卵巢发育的形态学与组织学研究[J].淮海工学院学报:自然科学版,2005,14(1):66-68.
[21] 杨玉香,梁维波,郑国富.辽东湾毛蚶繁殖季节研究[J].水产科学,2003,22(3):17-19.
[22] 王子臣,孙伯伦.渤海辽东湾毛蚶幼虫的数量变动和海区采苗[J].水产科学,1984,3(3):10-14.
[23] 竺俊全,杨万喜.毛蚶与青蚶精子超微结构及其所反映的蚶科进化关系[J].动物学研究,2004,25(1):57-62.
[24] 竺俊全.毛蚶Scapharca subcrenata精子发生过程中线粒体的行为研究[J].浙江海洋学院学报:自然科学版,2001,20(1): 23-26.
[25] 董迎辉,张晓,姚韩韩,等.毛蚶受精和早期卵裂过程核行为的荧光显微镜观察[J].台湾海峡,2011,30(2):223-228.
[26] 朱东丽,林志华,董迎辉,等.泥蚶(♀)×毛蚶(♂)受精及胚胎发育过程的初步研究[J].渔业科学进展,2010,31(5):69-75.
[27] 杜尚昆.毛蚶土池培养饵料育苗技术[J].齐鲁渔业,2008,25 (9):44-45.
[28] 梁维波,朱守维,杨玉香,等.毛蚶稚贝中间暂养技术的研究[J].齐鲁渔业,2008,25(3):30-31.
[29] 杨玉香,郭强,付卓.毛蚶室内人工育苗存在的问题及解决办法[J].齐鲁渔业,2009,26(1):42-43.
[30] 王丰,温尚明,杜尚昆.毛蚶人工升温育苗技术[J].河北渔业,2011(4):30-31.
[31] 葛立军,杨玉香,梁维波.不同饵料对毛蚶幼体发育的影响[J].水产科学,2008,27(5):226-229.
[32] 王友智,安宪深.毛蚶人工繁殖试验报告[J].河北水产科技, 1983(1):3-6.
[33] 刘爱英,马云聪,赵光环,等.毛蚶人工育苗技术[J].海洋湖沼通报,2005(1):86-90.
[34] 李永仁,郭永军,邢克智,等.北方毛蚶室内无附着基苗种繁育试验[J].科学养鱼,2014(5):42-43.
[35] 孙同秋,王玉清,王洪滨.黄河三角洲毛蚶大规格苗种池塘培育试验研究[J].齐鲁渔业,2011,28(5):15-17.
[36] 高霄龙,莉李,邱兆星,等.不同附着基对毛蚶幼虫附着变态影响的研究[J].海洋科学,2013,37(8):61-65.
[37] 徐国成,马甡,阎斌伦,等.影响毛蚶苗种培育的主要因子[J].水产科技情报,2007,34(5):195-198.
[38] 蒋宏雷,施祥元,刘伟健.毛蚶(Scapharaca subcrenata Lischke)人工育苗技术的初步研究[J].现代渔业信息,2006,21(8): 21-23.
[39] 沈伟良,尤仲杰,施祥元.饵料种类和密度对毛蚶浮游幼虫生长的影响[J].河北渔业,2007(9):18-20.
[40] 施祥元,沈伟良,尤仲杰.不同藻类对毛蚶浮游幼虫附着的影响[J].现代农业科技,2007(10):141-142.
[41] 沈伟良,尤仲杰,施祥元.温度与盐度对毛蚶受精卵孵化及幼虫生长的影响[J].海洋科学,2009,33(10):5-8.
[42] 顾晓英,尤仲杰,沈伟良,等.饵料对毛蚶(Scapharca subcrena+ ta)不同发育期幼虫脂肪酸组成的影响[J].海洋与湖沼, 2009,40(1):48-55.
[43] 杜尚昆.毛蚶幼体不同附着基附着效果试验[J].科学养鱼, 2008(4):23.
[44] 杜桂芝,杜尚昆.用300目沸石粉作为毛蚶稚贝附着基的方法[J].河北渔业,2013(6):30-31.
[45] 黄金臣,马云聪.利用室内水泥池中间培育毛蚶稚贝[J].河北渔业,2006(5):37-38.
[46] 葛立军,杨玉香,赵文,等.毛蚶稚贝池塘中间培育的最适密度研究[J].大连水产学院学报,2008,23(2):145-148.
[47] 王海涛,王世党,王同勇,等.毛蚶控温育苗与中间培育技术[J].中国水产,2010(2):38-39.
[48] 孙同秋,王玉清,王新叶,等.不同密度对毛蚶苗种生长影响的研究[J].齐鲁渔业,2010,27(1):14-16.
[49] 王友智,安宪深.毛蚶人工采苗试验[J].河北水产科技,1982 (6):19-21.
[50] 张建春,赵洪德.毛蚶半人工采苗及筏式笼养技术[J].河北渔业,2002(4):25-31.
[51] 谭围,陈傅晓,蒲利云,等.我国常见蚶类的人工育苗与养殖技术研究进展[J].现代农业科技,2012(8):327-330.
[52] 刘祖祥,朱善华.对虾和毛蚶的混养试验[J].海洋科学,1987 (6):56.
[53] 刘洪文.对虾与毛蚶池塘混养技术[J].河北渔业,2005(2): 22-41.
[54] 孙同秋,任贵如,王洪滨,等.黄河三角洲地区毛蚶高效生态养殖技术[J].齐鲁渔业,2010,27(9):23-25.
[55] 付卓,朱守维.锦州浅海毛蚶底播增养殖成效调查[J].河北渔业,2008(2):40-42.
[56] 战江祥,刘刚,王永安,等.海州湾毛蚶浅海筏式养殖试验[J].齐鲁渔业,2002,19(11):13.
[57] 林培振,刘德坤,朱志有.毛蚶筏式笼养技术[J].河北渔业, 2010(3):13-52.
[58] 高見東洋,吉岡貞範.サルボウガイ養殖試験[J].山口県内海水産試験場報告,1983,11:26-40.
[59] 圦本達也,那須博史,鳥羽瀬憲久,等.有明海たおいてカゴ飼育されたサルボウの成長,生残および摂餌状態[J].水産増殖,2007,55(4):535-540.
[60] 鈴木秀幸,山口啓子,瀬戸浩二.閉鎖性の高い中海で垂下養殖されたサルボウガイの成長と生残[J].水産増殖,2011, 59(1):89-99.
[61] 姜祖辉,王俊.温度和规格对毛蚶耗氧率和排氨率的影响[J].青岛大学学报:自然科学版,1999,12(1):77-81.
[62] 沈伟良,尤仲杰,施祥元.不同规格及不同盐度下毛蚶稚贝耗氧率和排氨率的研究[J].海洋水产研究,2008,29(2):53-56.
[63] 史宝,徐涛,马甡.盐度对毛蚶(Arca subcrenata Lischke)呼吸与代谢的影响[J].海洋湖沼通报,2008(1):104-108.
[64] 施祥元,尤仲杰,沈伟良,等.盐度对毛蚶稚贝生长和存活的影响[J].水产科学,2007,26(10):554-556.
[65] 鈴木秀幸,山口啓子,瀬戸浩二.中海たおけるサルボウガイの生残と成長たおよぼす低酸素および低塩分の影響[J].水産増殖,2012,60(2):261-268.
[66] 本田匡人,郡司掛博昭,松井繁明,等.サルボウガイ(Subcre+ nata kagoshimensis)の呼吸代謝た及ぼす低酸素の影響[J].九州大学大学院農学研究院学芸雑誌,2010,65(2):31-37.
[67] 方军,闫茂仓,张炯明,等.pH和氨氮对毛蚶稚贝生长与存活影响的初步研究[J].浙江海洋学院学报:自然科学版,2008, 27(3):281-285.
[68] 沈和定,张饮江,卢华春,等.毛蚶对单胞藻的滤除率、选择性和消化状况[J].上海水产大学学报,2006,15(2):195-200.
[69] 杨凤,刘丹,于倩,等.环境因子对毛蚶排遗和泥沙净化的影响[J].大连海洋大学学报,2012,27(6):523-527.
[70] 和田克彦,古丸明.サルボウガイの染色体核型[J].養殖研究所研究報告,1985(8):1-4.
[71] 陆荣茂,林志华,张永普,等.毛蚶、泥蚶、橄榄蚶的核型比较分析[J].上海水产大学学报,2008,17(5):625-629.
[72] 孙桂金,潘杰,刘可春,等.毛蚶、泥蚶和魁蚶ITS1核苷酸序列分析[J].生物技术通报,2011(7):117-120.
[73] 张波,孟学平.蚶科三种贝类ITS2核苷酸序列分析[J].石河子大学学报:自然科学版,2008,26(2):224-227.
[74] 郭永军,戴伟,王晓梅,等.毛蚶、魁蚶和泥蚶核糖体DNA转录间隔区的RFLP分析[J].华北农学报,2010,25(4):88-91.
[75] Tanaka T,Aranishi F.Mitochondrial DNA markers for PCR-based phylogenetic analysisof ark shells[J].Open Journal ofMarine Science,2013(3):182-189.
[76] 李旭光,阎斌伦,许广平,等.毛蚶三个地理群体生化遗传特征分析[J].海洋渔业,2007,29(3):207-213.
[77] 王晓梅,郭永军,戴伟,等.不同群体毛蚶rDNA转录间隔区RFLP及序列分析[J].四川动物,2008,27(5):774-776.
[78] 赵文,张雷,毕进红.辽宁沿海5个毛蚶群体遗传多样性的ISSR分析[J].水产学报,2011,35(6):854-862.
[79] 宋菲菲.山东近岸毛蚶群体的形态学和遗传学研究[D].青岛:中国海洋大学,2013.
[80] Tanaka T,Aranishi F.Genetic variability and population structure of ark shell in Japan[J].Open Journal of Marine Science,2014 (4):8-17.
[81] Feng Y W,Li Q,Kong L F.Isolation and characterization of 14 polymorphic microsatellite loci in the ark shell Scapharca subcre+ nata(Bivalvia:Arcidae)[J].Conservation Genetics,2009,10 (4):1125-1127.
[82] LiM,Zhu L,Zhou C Y,etal.Developmentand characterization of EST-SSRmarkers from Scapharca broughtonii and their transferability in Scapharca subcrenata and Tegillarca granosa[J].Molecules,2012,17(9):10716-10723.
[83] Dong Y H,Yao H H,Lin Z H,et al.Characterization of 62 polymorphic EST-SSR markers in the blood clam(Tegillarca grano+ sa)and their cross-amplification in Scapharca subcrenata[J]. Conservation Genetics Resources,2012(4):991-997.
A review:research progress of biology in ark shell Scapharca subcrenata insight from aquaculture
Abstract:The deterioration ofmarine environment and overexploitation lead to continual decline in stock of ark shell Scapharca subcrenata as an importantly economicmarinemollusk,and more attention has been paid to the resource recovery and aquaculture.The research progress on biological characteristics,reproduction cycle,hatching, culture,physiological ecology and genetics in the ark shell are summarized to provide some referential information for future study on resource enhancement viamarine ranching or releasing,artificial culture and genetic conservation.
Keywords:Scapharca subcrenata;biological property;artificial seeding;culture;genetics
中图分类号:S965;Q958.8
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2015.04.017
文章编号:2095-1388(2015)04-0437-07
收稿日期:2014-10-16
基金项目:国家自然科学基金资助项目 (31302186);辽宁省农业领域青年科技创新人才培养计划 (2014013);辽宁省海洋与渔业科学技术计划项目 (201209,201301);辽宁省科学技术计划项目 (2014203006)
关键词:毛蚶;生物学特性;人工繁育;人工增养殖;遗传学