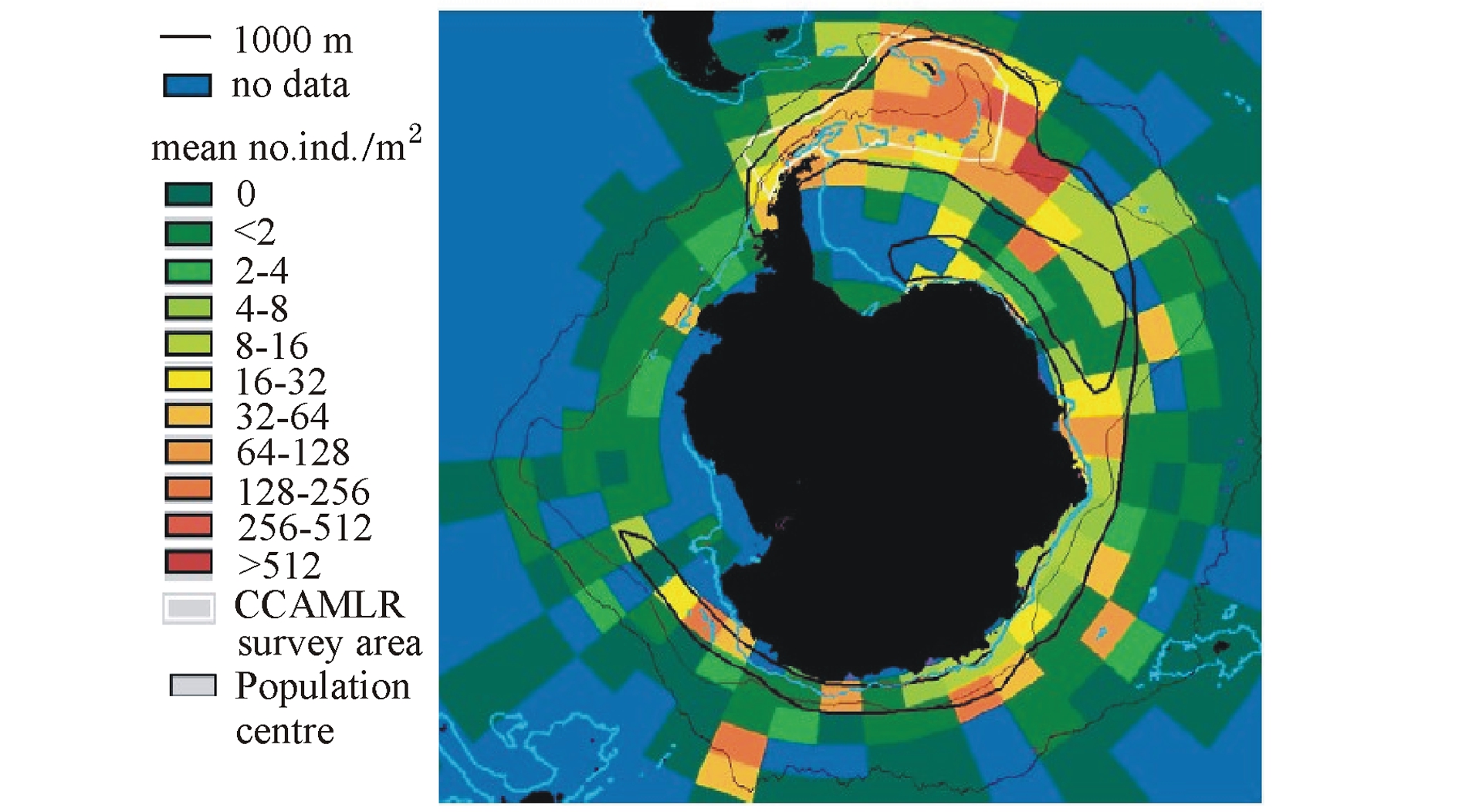
图1 南极磷虾密度的环极分布图(Atkinson等[11])
Fig.1 Circumpolar distribution of krill density
摘要:环境因素是影响南极磷虾Euphausiasuperba分布的关键因素,是研究南极磷虾渔场形成机理及渔情、渔汛预报的基础。为了准确评估磷虾的资源量,扩大对磷虾资源的开发规模,使磷虾能够成为可持续开发的生物资源,国内外学者持续开展了南极磷虾的资源调查,对磷虾的分类、资源量、分布、渔场和部分相关环境因素进行了研究,并取得了显著的进展。本研究中主要综述了环境因素,包括非生物环境因素和生物环境因素,对南极磷虾资源分布影响的研究进展,并对研究中提出的一些假说做了阐释,以期为今后环境因素对南极磷虾资源分布影响的进一步研究提供参考。
关键词:南极磷虾;生物资源;环境因素;资源量;资源分布
南极磷虾Euphausiasuperba是南大洋生态系统中的关键生物,很有可能成为人类可以利用的最大的海洋蛋白资源,被誉为是 “人类未来的动物蛋白库”[1]。尽管南极比较遥远而且环境恶劣,但南极磷虾巨大的生物量及利用价值深深地吸引着人类,使它在很早就成为一个商业开发的捕捞对象。国际上对南极磷虾的研究开展得比较早,但早期主要以生物学研究为主。随着各国对南大洋战略地位的重视和科研投入的增加,目前对南极磷虾的研究已成为研究南极的重点和热点。为了建立合适的养护管理措施,近年来的研究主要集中在磷虾种群资源的丰度及分布方面[2]。调查范围已覆盖南大洋的大部分海域,其中包括南极半岛和西部的斯科舍海[3],以及印度海域[4]和西部的罗斯海[5]。资源的开发必须建立在准确评估的基础之上,而环境因素作为影响海洋生物资源的重要因素,对海洋生物资源的准确评估影响显著。南大洋的海洋环境较为复杂,环境因素对南极磷虾丰度及分布有显著影响[6], 如海冰范围、盐度、水温和环流方式, 这些因素的综合效应使得监测和评估磷虾资源状况变得非常困难[7], 目前尚未有研究结果能够清楚地表明环境因素与磷虾分布之间的关系。但是,研究环境因素对南极磷虾资源分布的影响,不仅可以深入了解磷虾资源的数量和分析磷虾渔场的形成机制,同时也可掌握磷虾补充量及作业渔场的分布情况[8],对南极磷虾资源的可持续开发具有指导意义。
每一种生物都有其特定的生活地域,南极磷虾主要分布于南大洋的各个水域,是由南大洋海域的地理环境因素决定的。南大洋位于南极洲周围的太平洋、大西洋、印度洋和附属海域的南部。海底地貌特征包括海底高原,隆起海盆,水下深度不超过2000 m,为一相当平坦的地区,常覆盖很厚的沉积物。南极磷虾的分布区位于50°S以南水域,呈环南极的分布形态,密集区常出现在陆架边缘、冰架边缘和岛屿周围[9-11](图1)。Atkinson等[11]认为,南极磷虾未成熟个体主要分布在大陆架及边缘,而成熟个体主要分布在大陆架和大陆坡水域,产卵个体主要分布在大陆坡至大陆隆。在冬季磷虾的丰度比较低,主要分布在大陆架附近,从11月起磷虾的丰度快速增加,沿着大陆架坡折向北部大洋水域并且延伸直至极限59°S海域,在1—2月磷虾在最北部的分布丰度达到最大(图2)。由此可见,随着季节的变化,磷虾的分布会沿着大陆架的延伸做来回的迁移。另外,磷虾白天主要分布在较深水层,夜晚才会浮到上层水域进行摄食等。对于磷虾分布的这种形式学者们提出了多方面的解释,包括海冰、海流、捕食者和食物等[12-16],但任何一个方面都不能对这一现象做出充分解释,因为每一个方面都有例外的情况存在[8]。有时一年中磷虾的资源量与环境之间的关系很明显,但这种关系在年际间及不同地域间并不连续[2]。在小范围区域内,磷虾与环境之间的关系中也只有磷虾密度与水深的关系在年际间连续[10]。
对南极磷虾资源分布具有较大影响的主要环境因素分为非生物环境因素和生物环境因素[17],其中非生物环境因素主要包括海洋地理、海洋水文动力和海水理化因素,生物环境因素则主要包括浮游动植物等。此外,关于海洋地理因素对南极磷虾分布影响的研究文献较少,因此,本研究中着重讨论其他几种环境因素的作用。
2.1海洋水文动力因素
2.1.1 海冰 在复杂的南大洋海洋环境中,海冰对南极磷虾资源分布的影响尤其重要[18]。海冰作为南极磷虾栖息地, 为正在越冬的南极磷虾成体和幼体提供了很好的饵料环境[13]。在冬末,南极磷虾主要栖息在浮冰下并从浮冰底部获取冰藻作为食物来源[19-20]。因此,冬、春季海冰范围不仅对磷虾成年体及未成熟个体的生长产生影响[19,21],而且也对磷虾补充量及作业渔场的分布具有重要的影响[8]。48海区是南极磷虾主要的分布区域,也是捕捞活动最密集的区域,因此,针对该海区海冰与南极磷虾分布关系的研究较多。有研究表明:48海区南极磷虾资源夏季丰度与上一年度冬季海冰的面积成正比[22-23],在捕捞开发中磷虾CPUE年间的变化趋势也是随着海冰密集度的增加而变大的[24]。此外,陈峰等[25]针对48海区利用 1997—2008 年南极磷虾产量及各月南极海冰面积数据, 分析了南极磷虾产量的时空分布, 以及海冰对南极磷虾资源丰度的影响,进一步证实了南极磷虾的分布随海冰面积的变化关系。近年来,由于受地球变暖的影响,使得在南极磷虾作为躲避捕食者和保护自身中起着重要作用的海面浮冰在减少,致使不少南极磷虾因失去了掩盖体而被捕食,所以部分水域的南极磷虾个体数在锐减[26]。南极的海冰对极地其他生物种群以及气候、海洋、冰和南极食物网之间的联系等也具有重要作用,其分布范围、密集度等都可能对当地初级生产力、海豹捕食区域和企鹅的活动范围等产生影响。因此,对南极海冰的监测、研究和预报具有重要的现实意义和长远意义。
2.1.2 海流和水团 南大洋环流系统一般可分为南极绕极流ACC(the antarctic circumpolar current)和沿岸流两部分。海水按温盐结构可分为5个水团:南极陆架水、南极底层水、绕极深层水、南极表层水和南极中层水。这些海流及水团对南极磷虾资源的分布及渔场的形成均有明显的影响和作用[27,4]。海水的垂直交换不仅带来了大量的硝酸盐、磷酸盐等营养物质, 促使浮游生物大量繁殖, 为南极磷虾提供了丰富的饵料[11],而且有利于维持适宜的海表温度。此外,分布在大陆架斜坡海域的磷虾种群, 被认为是南极绕极流输送的产物[3],Lascar等[28]认为,南极绕极流对南极磷虾幼体有很强的输送作用,但是还没有证据能证明磷虾只是被动地随海流漂游。环流方式可能也对磷虾资源的分布产生影响, 图3为南极主要海流的环流形式,对照该图与南极磷虾环南极的密度分布图(图1)可以看出,南极磷虾种群的分布与主要的海流分布整体上是一致的[11]。对于较大规模或是中等规模的虾群,海流对其分布都具有重要的影响,而且有关学者对此基本能够达成共识。但是海流与磷虾资源分布之间可能并不是简单的因果关系,影响南极磷虾资源丰度的环境因素是多方面的[29],磷虾是随海流漂移还是在进行洄游尚且没有直接的证据予以说明,故不能简单地认为磷虾的分布与海流分布相一致是海流运输的结果。因此,海流和水团对南极磷虾的分布、洄游、繁殖与生长的影响机制还需进一步研究,分析海流、水团的分布及其变动也是研究南极磷虾资源时空动态及渔场形成机理的重要内容。
2.2海水理化因素
2.2.1 水温 南极磷虾是一种喜冷水环境的浮游动物,成体栖息于200 m以浅水域,幼体则在表层水域中栖息的时间较长,其对海洋温度和海冰密度极为敏感。磷虾栖息地分布范围的极限温差约为7 ℃,1~2 ℃的水温变化对南极磷虾生理功能、分布和行为会产生显著影响。因此,水温是影响南极磷虾分布的一个重要因素[30]。目前,已有试验证明,温度对磷虾的生长具有重要的影响,磷虾的蜕壳周期会随温度的变化而改变[31-32]。海表温度(SST)作为一个非常重要的海洋要素,也是控制生物种群分布及其洄游和繁殖过程的基本环境参量,在现行海洋渔场渔情分析预报中占有重要地位。南极地区的温跃层变化很小,通常表层和底层温差仅1 ℃左右[33],所以SST的季节变化仅考虑水平的变化。试验证明,磷虾短期内能够忍耐水温出现一定的波动,但其新陈代谢需求及肌体组织无法维持较大的水温变化。由此推断,南极磷虾资源与SST之间必然存在着密切的关系[34]。Trathan等[10]利用GAM模型对夏季开始时南乔治亚群岛水域南极磷虾体长指数与SST之间的关系进行了初步分析,发现在SST较高的几个年份内磷虾的补充量较低,认为南乔治亚群岛夏季南极磷虾丰度与温度之间存在负相关关系。朱国平等[35]利用GAM模型分析了SST与南极磷虾CPUE的分布关系,结果表明:南奥克尼群岛水域南极磷虾作业渔场的适宜海水表温为0.1~1.8 ℃,最适海水温度为0.5~1.5 ℃。Vanyushin[36]认为,夏季渔汛初期的SST信息可用于预测整年的南极磷虾产量。由于南大洋水域面积广阔,且磷虾资源分布较为广泛,所以水温主要是通过影响磷虾个体的生长间接地对磷虾资源量的分布产生一定的影响[37]。
2.2.2 光照 光照作为一个最基本的环境因子,对地球上的一切生物来说都是必不可少的。南极日照随着季节的变化具有很大的差异,而磷虾作为南极复杂环境的适应者,其行为分布应该是与南极的光照变化相一致,但至今还没有学者将南极磷虾资源的分布与光照周期的变化建立直接的联系。目前,对光照与南极磷虾关系的研究仅局限于南极磷虾的行为和生理方面。Strand 等[38]在实验水槽中观察化学和视觉刺激对磷虾集群行为的影响时发现,在2.12 lx光照强度下,南极磷虾集群的速度最快,光照强度小幅变化或者渐变对集群没有影响,且当光照强度不高时,集群能够迅速恢复。Teschke 等[39-40]在实验室环境下依照南大洋夏季、秋季和冬季不同条件的光照周期模拟不同的光照密度,对南极磷虾进行模拟日照试验,然后分析光照对磷虾的进食、新陈代谢速率性和成熟度的影响。结果表明,不同的光照环境对于磷虾的生理生长和成熟能产生周期性的重要影响,并指出了光照这一环境因素在磷虾生存史上的重要意义。Sear等[41]针对不同光照周期对磷虾基因表达的影响进行了研究,从分子学的角度分析了在不同的光照周期下,基因表达的变化对磷虾的新陈代谢、肌动活动、蛋白质合成,以及其他各种细胞活动的影响,证实了不同的光照周期对南极磷虾控制蜕壳、摄食和游动的基因的表达具有明显的影响。
浮游生物是海洋鱼类饵料的基础,也是渔场形成的根本条件[42]。南极磷虾主要以水体中的浮游植物(如硅藻等)为食物,但是当食物比较匮乏时,南极磷虾的摄食习性也会发生改变,会通过摄食樽海鞘等浮游动物来度过食物匮乏时期[43]。除此之外,在自然条件下,有些樽海鞘类的肠胃中经常会含有磷虾幼体,因此,樽海鞘类和南极磷虾在摄食浮游植物方面属于竞争关系[44]。樽海鞘类和南极磷虾的这种生态关系可能也是南大洋浮游动物生态系统研究的重要方面。南极夏季水体中的浮游植物繁殖速度很快,初级生产力非常高,为南极磷虾提供了充足的食物和能量。叶绿素浓度是海洋浮游植物以及海洋动力过程的重要指标。通过获取海水中叶绿素的信息便可以判断海水浮游植物含量的高低及其变化,然后进行渔场分析和渔业资源评估。Marrari等[45-46]研究了1997—2004年南极半岛西侧海域(55°~75°S、50°~80°W)叶绿素a的分布,通过对比分析南半球夏季叶绿素a的分布(图4)与磷虾的分布(图1),认为在南半球夏季,磷虾分布同叶绿素分布高度相关。从空间上来看, 在其栖息水域范围内, 夏季南极磷虾的资源密度与叶绿色a 浓度呈正相关关系[47],而这只能表明夏季食物来源是影响部分海域南极磷虾密度的关键因素。南极磷虾资源大范围的分布与叶绿素的密度分布为非线性关系[11],尽管当藻类暴发时,磷虾可获得最大的生长速率[48],但这并不能说明磷虾需要浮游植物的暴发来进行生长。实际上,无论是在夏季还是冬季,磷虾都能够很好地适应食物的匮乏,比如它可以通过改变摄食对象甚至是“负生长”来维持生命。 有效地摄食也许是磷虾在南大洋中成功生存的关键,但该生存机制还需进一步阐明。
南极磷虾作为南大洋生态系统中的关键种和生物资源量最大的物种,在整个南大洋的生态系统中十分重要。此外,极地变暖正在逐渐加剧[26],而这些海域的中上层无脊椎动物大多为狭温动物,它们对环境的变化非常敏感。而磷虾作为在极地成功生存的典型生物,尽快掌握其生存机制,才能更好地预测其未来的情况。随着全球气候的变化及人类参与的捕捞活动,南极磷虾的数量正在逐年减少[26]。为了使磷虾能够成为可持续开发的生物资源,进一步了解环境因子对磷虾资源的影响迫在眉睫。目前的研究还难以表明磷虾跟环境因素的明确关系,大部分研究只是尝试着建立海冰、海流和叶绿素与磷虾资源分布的关系,还不能以此研究磷虾在南极成功生存的机制。温度、盐度、敌对生物等对南极磷虾群分布的影响,以及纬度、海水的密度与稳定性和有机物的溶解及其营养富集浓度与磷虾密度的相关性研究还很少涉及。
图1 南极磷虾密度的环极分布图(Atkinson等[11])
Fig.1 Circumpolar distribution of krill density
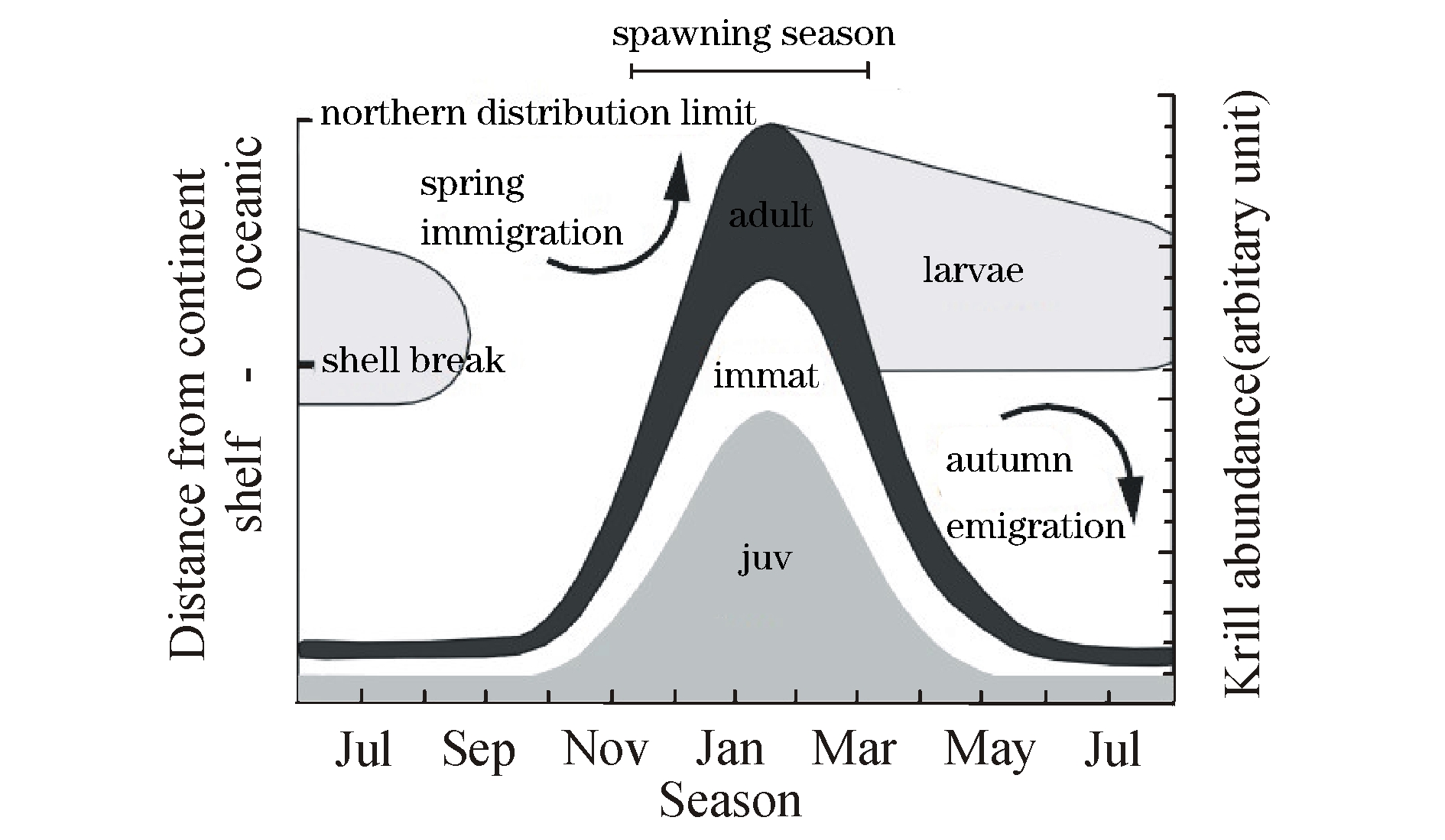
图2 南极磷虾成虾、未成熟虾和幼虾和产卵个体资源丰度分布随季节变化图(Siegel[2])
Fig.2 The changes in abundance distribution of adults,immature and juvenile krill at various seasons
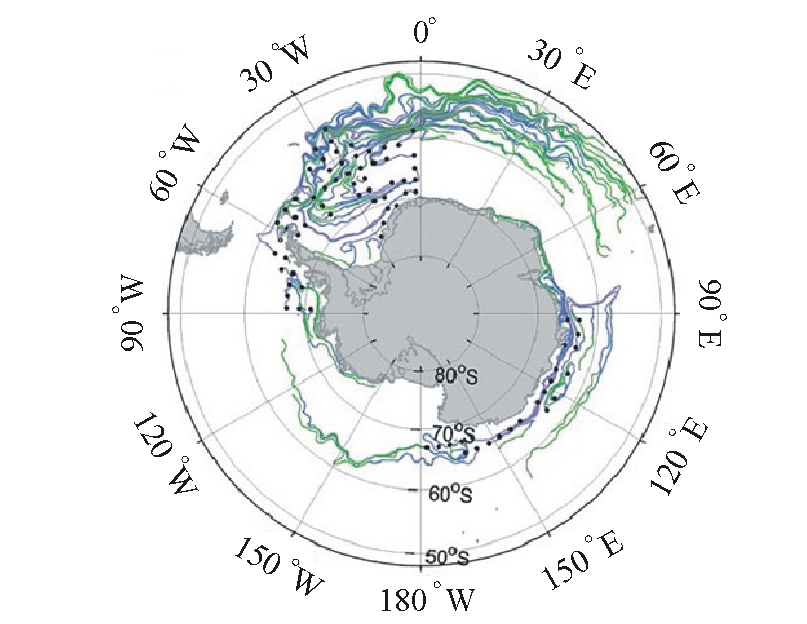
图3 南极海流的分布图
Fig.3 The distribution of large scale advection in Antarctic Ocean(Atkinson等[11])

图4 基于7个季节(1997—2004年南半球夏季)SeaWiFS数据的南大洋开放水域季节中月均叶绿素密度分布值(Atkinson等[11])
Fig.4 Monthly mean Chl a values in the main open water seasons,based on 7 seasons of SeaWiFS data (from summers of 1997 to 2004)
盐度作为海水的特性之一,对水生物机体的生理反应影响较大,不同盐度下的水生生物会表现出不同的适应状态[49]。南大洋被认为是地球上最大的高硝酸盐和低叶绿素(HNLC)海区[50], Martin等[51]提出,在HNLC海区铁是浮游植物生产力的限制因子,为此各国的海洋学家进行了大量的铁加富实验,并初步确认了铁对浮游生物的限制作用[52]。而且在极锋区附近,底层流与大陆架沉积物接触可引起溶解铁浓度的升高[53],在靠近海岛或大陆架的海区,表层溶解铁浓度也相对较高[54-56]。这与海水叶绿素和磷虾的分布较为一致。除此之外,磷虾的生长需要不断的蜕壳,食物匮乏时磷虾还可以通过蜕壳进行“负生长”来维持生命。在蜕壳过程中磷虾需要与外界环境进行一系列物质循环与能量转换,这不仅需要适宜的盐度来维持渗透压[57],而且需要从外界摄取大量的无机盐如钙、磷、铁、硅等来生成新的外壳。另外,南极磷虾的富氟异常,其含量是海水中氟含量的3000倍,南极磷虾对氟的富集不仅富集率高,且速度极快[58-59],因此,海水中氟的含量对磷虾的生长应该也有很大影响。由此推断,盐度可以通过两个方面来影响磷虾的资源量和分布,一个是通过影响磷虾自身的生理生长,另一个则是通过影响磷虾所摄食的浮游生物的生长。但至今国际上还没有人验证出盐度与磷虾资源分布的关系。
由美、英、德、澳等国和国际捕鲸委员会(International whaling commission,IWC)联合实施的南大洋生态系统动力学(SO-GLOBEC)项目,正尝试着从生态系统水平上研究磷虾与其竞食者和捕食者之间的关系[60]。目前,对南极磷虾的摄食能力和躲避被捕能力之间的相对关系的研究还比较少。在磷虾的种群动力学中,死亡率对其数量的控制无疑是最主要的方面[61],每年都有超过一亿t的磷虾被捕食者消耗掉[62]。捕食者在靠近岛屿的地方繁殖时,捕食活动更加剧烈[63]。鲸鱼等大型捕食者的减少导致磷虾过剩的现象更反映了捕食者对磷虾资源量变动的控制[12]。此外,捕食者还对磷虾的行为具有很大的影响,比如,磷虾的集群行为可以增加躲避捕食者的优势。有观察发现,在秋末和冬季磷虾的集群行为比较活跃,而且当有捕食者存在时,磷虾的集群更加密集;当不存在捕食者时,磷虾的集群就没那么密集[64]。Zhou等[65]对磷虾的集群与垂直移动活动进行了研究,指出磷虾昼夜垂直移动的习性也与躲避捕食者有关。因此,捕食者对磷虾的资源量和分布具有一定的影响。Atkinson等[11]在南极磷虾的环极海洋栖息地的综述中,通过分析磷虾的生命史及主要环境因素与磷虾分布的关系,提出了磷虾最适栖息地的“风险补偿”模型假说,根据该假说,磷虾不选择浮游植物密度最大的区域作为主要栖息地,主要是因为在这些区域虽然食物丰富,但捕食者的分布密度也比较大,因此被捕食的风险也大,为了躲避捕食者,磷虾群体主要集中分布在浮游生物密度适中、被捕食风险较小的区域,并且通过白天下沉至深海晚上浮到水面摄食的习性来减小被捕食的风险。虽然从以往数据的类比分析中可以进行如此推断,但由于缺少捕食者及其他各环境因子与磷虾分布的连续数据,目前对该假说还停留在推理阶段。
南极海域广阔,环境复杂,数据的采集比较困难,且中国对南极磷虾资源的调查时间较短,掌握的数据很少,很难系统地做出环境因子对磷虾资源分布的影响分析。目前,中国对南极磷虾资源的研究还主要集中在对其的捕捞开发和加工利用上,环境因素对磷虾资源的影响研究还比较少,也比较局限,这对今后磷虾资源的研究分析是不利的,也将会影响中国对磷虾资源进行长期开发的效率。随着科学技术的不断发展,用于渔业资源探测及环境调查的仪器不断更新,相关数据的采集将不再那么困难。在今后的调查研究中,对于相关环境数据的采集要做到更加全面、系统和准确。作者认为,磷虾的分布形式是磷虾的行为和海洋物理、化学、生物环境综合作用的结果,掌握磷虾的行为对于磷虾分布形式的作用是了解南大洋生态系统的基础。因此,在今后可以针对磷虾的个体行为和群体行为展开深入研究,比如海流、盐度和海冰对磷虾生长和行为的影响等。此外,南极磷虾的行为和资源量还与南大洋生态系统中的哺乳动物、鱼类、鸟类和其他浮游动物有很大关系,在对南极磷虾资源量和分布的调查研究中,应结合南大洋的其他鱼类、哺乳动物和鸟类的分布情况,尝试在基础研究上综合各个环境因子,从整个生态系统的角度,建立磷虾的海洋生态系统动力学模式对磷虾的种群进行分析,这才有可能更加清楚地掌握磷虾资源和分布的变动规律。要确切地掌握环境因素对磷虾资源分布的影响,还需要各国之间的团结协作,数据共享,长期努力,早日实现磷虾资源的可持续开发。
参考文献:
[1] 孙松.南极磷虾[J].世界科技研究与发展,2002,24(4):57-60.
[2] Siegel V.Distribution and population dynamics ofEuphausiasuperba:summary of recent findings[J].Polar Biol,2005,29:1-22.
[3] Hofmann E E,Murphy E J.Advection,krill,and Antarctic marine ecosystems[J].Antarct Sci,2004,16:487-499.
[4] Nicol S.Krill and currents—physical and biological interactions influencing the distribution ofEuphausiasuperba[J].Ocean Polar Res,2003,25:633-644.
[5] Faranda F M,Guglielmo L,Anora A.Ross Sea Ecology[M].New York:Springer,2000.
[6] 樊伟,伍玉梅,陈雪忠,等.南极磷虾的时空分布及遥感环境监测研究进展[J].海洋渔业,2010,32(1):95-101.
[7] Smetacek V,Nicol S.Polar ocean ecosystems in a changing world[J].Nature,2005,437:362-368.
[8] Siegel V,Loeb V.Recruitment of Antarctic krillEuphausiasuperbaand possible causes for its variability[J].Marine Ecology Progress Series,1995,123:45-56.
[9] Carin J A,Cabell S,Davis S M,et al.Distribution of larval krill and zooplankton in association with hydrography in Marguerite Bay,Antarctic Peninsula,in Austral fall and winter 2001 described using the Video Plankton Recorder[J].Deep-Sea Research Ⅱ,2008,55:455-471.
[10] Trathan P N,Brierley A S,Brandon M A,et al.Oceanographic variability and changes in Antarctic krill (Euphausiasuperba) abundance at South Georgia[J].Fish Oceanogr,2003,12(6):569-583.
[11] Atkinson A,Siegel V,Pakhomov E,et al.Oceanic circumpolar habitats of Antarctic krill[J].Marine Ecology Progress Series,2008,362:1-23.
[12] Mackintosh N A.Distribution of post-larval krill in the Antarctic[J].Discovery Rep,1973,36:95-156.
[13] Brierley A S,Fernandes P G,Brandon M A,et al.Antarctic krill under sea ice:elevated abundance in a narrow band just south of the ice edge[J].Science,2002,295:1890-1892.
[14] Marr J W S.The natural history and geography of the Antarctic krill (EuphausiasuperbaDana)[J].Discovery Rep,1962,32:433-464.
[15] Pakhomov E A.Demography and life cycle of Antarctic krill,Euphausiasuperba,in the Indian sector of the Southern Ocean:long-term comparison between coastal and open ocean regions[J].Can J Fish Aquat Sci,2000,57:68-90.
[16] Nicol S.Krill,currents,and sea ice:Euphausiasuperbaand its changing environment[J].Bioscience,2006,56:111-120.
[17] Lawson G L,Wiebe P H,Stanton T K,et al.Euphausid distribution along the Western Antarctic Peninsula-Part B:distribution of euphausid aggregations and biomass,and associations with environmental features[J].Deep-Sea Research Ⅱ,2008,55:432-454.
[18] Mackintosh N A.Life cycle of Antarctic krill in relation to ice and water conditions[J].Discovery Rep,1972,36:1-94.
[19] Marschall H P.The overwintering strategy of Antarctic krill under the pack-ice of the Weddell Sea[J].Polar Biology,1988,9:129-135.
[20] Stretch J J,Hamner P P,Hamner W M,et al.Foraging behavior of Antarctic krillEuphausiasuperbaon sea ice microalgae[J].Mar Ecol Prog Ser,1988,44:131-139.
[21] Siegel V,Loeb V,Groger J.Krill (Euphausiasuperba) density,proportional and absolute recruitment and biomass in the Elephant Island region (Antarctic Peninsula) during the period 1977 to 1997[J].Polar Biology,1998,19:393-398.
[22] Hewitt R P,Demer D A,Emery J H.An 8-year cycle in krill biomass density inferred from acoustic surveys conducted in the vicinity of the South Shetland Islands during the Austral summers of 1991-1992 through 2001-2002[J].Aquatic Living Resources,2003,16(3):205-213.
[23] Atkinson A,Siegel V,Pakhomov E,et al.Long-term decline in krill stock and increase in salps within the Southern Ocean[J].Nature,2004,432:100-103.
[24] 戴立峰,张胜茂,樊伟.南极磷虾资源丰度变化与海冰和表温的关系[J].极地研究,2012,24(4):352-359.
[25] 陈峰,陈新军,刘必林,等.海冰对南极磷虾(Euphausiasuperba)资源丰度的影响[J].海洋与湖沼,2011,42(4):495-499.
[26] 缪圣赐.因受地球变暖的影响南极磷虾的个体数在锐减[J].现代渔业信息,2008,23(7):33.
[27] Hofmann E E,Klinck J M,Locarnini R A,et al.Krill transport in the Scotia Sea and environs[J].Antarct Sci,1998,10:406-415.
[28] Lascar A C M,Hofmann E,Ross R M,et al.Seasonal variability in the distribution of Antarctic krill,Euphausiasuperba, west of the Antarctic Peninsula[J].Deep-Sea Research I,1999,46:951-984.
[29] Siegel V.A concept of seasonal variation of krill (Euphausiasuperba) distribution and abundance west of the Antarctic Peninsula[C]//Sahrhage D.Antarctic Ocean and resources variability.New York:Springer,1988:219-230.
[30] 朱国平,朱晓艳,孟涛,等.2012年秋冬季利文斯顿岛南极磷虾渔业CPUE指标变动及其影响因素[J].大连海洋大学学报,2013,28(4):399-405.
[31] Maihara Y,Endo Y.Laboratory observations on molting and growth of Antarctic krillEuphausiasuperbaDana[J].Mem Nat Inst Polar Res Special,1986(1):125-127.
[32] Nicol S,Stolp M A.Refinement of the moult staging technique for Antarctic krill(Euphausiasuperba)[J].Mar Biol,1990,104:169-173.
[33] Arrigo K R,Worthen D,Schnell A,et al.Primary production in Southern Ocean waters[J].Journal of Geophysical Research,1998,103:15587-15600.
[34] Huang T,Sun L,Stark J,et al.Relative changes in krill abundance inferred from Antarctic fur seal[J].PLoS One,2011,6(11):e27331
[35] 朱国平,冯春雷,吴强,等.南极磷虾调查CPUE指数变动的影响因素初步分析[J].海洋渔业,2010,32(4):368-373.
[36] Vanyushin G P.Sea-surface temperature and krill catches around South Georgia in December-February 1989-1991 and 1999-2001[J].CCAMLR Science,2002,9:165-172.
[37] Brown M,Kawaguchi S,Candy S,et al.Temperature effects on the growth and maturation of Antarctic krill (Euphausiasuperba)[J].Deep-Sea Research Ⅱ,2010,57:672-682.
[38] Strand S W,Hammer W M.Schooling behavior of Antarctic krill (Euphausiasuperba) in laboratory aquaria:reactions to chemical and visual stimuli[J].Marine Biology,1990,106:355-359.
[39] Teschke M,Kawaguchi S,Meyer B.Effects of simulated light regimes on maturity and body composition of Antarctic krill,Euphausiasuperba[J].Marine Biology,2008,154(2):315-324.
[40] Teschke M,Kawaguchi S,Meyer B.Simulated light regimes affect feeding and metabolism of Antarctic krill,Euphausiasuperba[J].Limnol Oceanogr,2007,52:1046-1054.
[41] Sear P,Geraint A T,Teschke M,et al.Effects of simulated light regimes on gene expression in Antarctic krill (EuphausiasuperbaDana)[J].Journal of Experimental Marine Biology and Ecology,2009,38:157-164.
[42] 牛明香,李显森,徐玉成.智利外海竹筴鱼中心渔场时空变动的初步研究[J].海洋科学,2009,33(11):105-109.
[43] Kawaguehi S,Takahashi Y.Antarctic krill (EuphausiasuperbaDana) eat salps[J].Polar Biol,1996,16:479-481.
[44] Kawamura A,Michimori K,Moto J.Marked inverse distribution of salps to other macrozooplankton in waters adjacent to the South Shetland Islands[J].Polar Biol,1994,7:70-81.
[45] Marrari M, Daly K L,Hu C M.Spatial and temporal variability of Sea WiFS chlorophyll a distributions west of the Antarctic Peninsula:Implications for krill production[J].Deep-Sea Research Ⅱ,2008,55:377-392.
[46] Marrari M, Hu C M,Daly K L.Validation of Sea WiFS chlorophyll a concentrations in the Southern Ocean:a revisit[J].Remote Sensing of Environment,2006,105:367-375.
[47] 朱国平.基于广义可加模型研究时间和环境因子对南极半岛北部南极磷虾渔场的影响[J].水产学报,2012,36(12):1863-1871.
[48] Ross R M,Quetin L N,Baker K S,et al.Growth limitation in youngEuphausiasuperbaunder field conditions[J].Limonol Oceanoger,2000,45(1):31-34.
[49] Fry F E J.The effect of environmental factors on the physiology of fish[M]//Hoar W S,Randall D J.Fish Physiology Vol Ⅵ:Environmental Relations and Behavior.New York:Academic Press,1971:1-98.
[50] Watson A.Iron limitation in the oceans[C]//The Biogeochemistry of Iron in Seawater.Chichester,UK:John Wiley and Sons,2001:9-39.
[51] Martin J H,Fitzwater S E.Iron-deficiency limits phytoplankton growth in the northeast Pacific Subarctic[J].Nature,1988,331:341-343.
[52] 孙松,蒲新明,张永山.南大洋普里兹湾的铁加富实验:对铁假说的检验[J].中国科学D辑:地球科学,2009,39(2):212-221.
[53] Löscher B W,Baar H J W,Jong J T M,et al.The distribution of Fe in the Antarctic Circumpolar Current[J].Deep-Sea Res Ⅱ,1997,44:143-187.
[54] Martin J H,Coale K H,Johnson K S,et al.Testing the iron hypothesis in ecosystems of the equatorial Pacific Ocean[J].Nature,1994:123-129.
[55] Planquette H,Statham P J,Fones G R,et al.Dissolved iron in the vicinity of the Crozet Islands,Southern Ocean[J].Deep-Sea Research Ⅱ,2007,54:1999-2019.
[56] Fitzwater S E,Johnson K E,Gordon R M,et al.Trace metal concentrations in the Ross Sea and their relationships with nutrients and phytoplankton growth[J].Deep-Sea Research Ⅱ,2000,47:3159-3179.
[57] 李二超.盐度对凡纳滨对虾的生理影响及其营养调节[D].上海:华东师范大学,2008.
[58] Soevik T,Braekkan O R.Fluoride in Antarctic krill (Euphausiasuperba) and Atlantic krill (Meganyctiphanesnoroegica)[J].Fish Res Board Can,1979,36:1414-1416.
[59] 张海生.南大洋氟的生物地球化学研究Ⅱ.氟在南极磷虾甲壳中的动态变异及其富集原因[J].极地研究,1992,4(1):17-22.
[60] Robin M R,Langdon B Q,Timothy N,et al.Growth and behavior of larval krill (Euphausiasuperba) under the ice in late winter 2001 west of the Antarctic Peninsula[J].Deep-Sea Research Ⅱ,2004,51:2169-2184.
[61] Murphy E J,Reid K.Modeling Southern Ocean krill population dynamics:biological processes generating fluctuations in the South Georgia ecosystem[J].Mar Ecol Prog Ser,2001,217:175-189.
[62] Mori M,Butterworth D S.A first step towards modeling the krill-predator dynamics of the Antarctic ecosystem[J].CCAMLR Sci,2006,13:217-277.
[63] Fraser W R,Hofmann E E.A predator’s perspective on causal links between climate change, physical forcing and ecosystem response[J].Mar Ecol Prog Ser,2003,265:1-15.
[64] Ainley D G,Ballard G,Dugger K M.Competition among penguins and cetaceans reveal trophic cascades in the Western Ross Sea,Antarctica[J].Ecology,2006,87:2080-2093.
[65] Zhou M,Dorland R D.Aggregation and vertical migration behavior ofEuphausiasuperba[J].Deep-Sea Research Ⅱ:Topical Studies in Oceanography,2004,51:2119-2137.
Abstract:Environmental factors are the key factors that have great effect on distribution of Antarctic krillEuphausiasuperba, and the research foundation of formation mechanism of fishing grounds, fishing conditions and fishing season forecast. For decades, considerable efforts have been devoted and significant progress has been made to understanding krill life histories and the interaction of krill populations with regional circulation and other environmental factors in order to make an accurate assessment of the biomass of krill and to expand the scale of the exploration of krill resource. This view describes primarily the current status of the research expounding the relationships between krill distribution and environmental factors, including abiotic and biological environmental factors, intending to indicate what studies are available and where additional work is needed. The further explanations are mad for some hypothesis to provide the reference to further research on the impact of environmental factors on the Antarctic krill resources distribution.
Key words:Euphausiasuperba; biology resource; environmental factor; biomass; resource distribution
DOI:10.3969/J.ISSN.2095-1388.2014.03.024
文章编号:2095-1388(2014)03-0316-07
收稿日期:2013-09-04
基金项目:南极海洋生物资源开发与利用项目(D-8003-09-0158);上海市重点学科建设项目(S30702)
中图分类号:S932.4
文献标志码::A