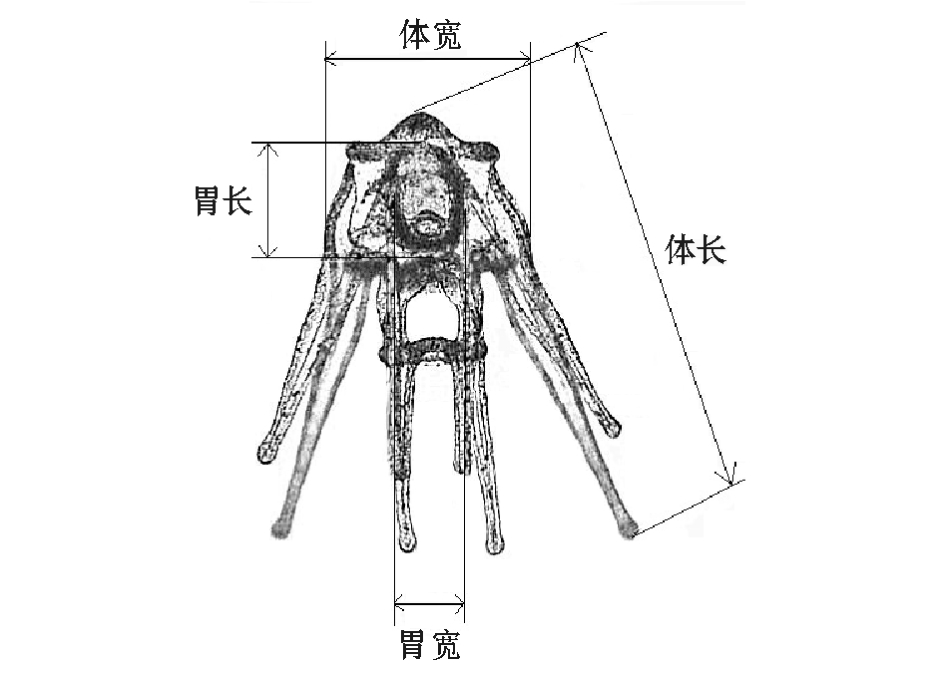
图1 海胆幼体体长、体宽、胃长、胃宽的测量
Fig.1 M easurement of body length,body w idth, stomach length and stom ach w id th in sea urchin larvae
摘要:通过构建不同近交水平的海胆群组,即半同胞近交组 (H组,F=0.125)、全同胞近交组 (F组,F=0.25)和对照组 (C组,F=0),研究了近交对中间球海胆Strongylocentrotus intermedius受精、孵化和幼体发育阶段的影响。结果表明:近交对海胆的受精率、孵化率和四腕幼体的畸形率均无显著影响 (P>0.05);不同近交水平的四腕、六腕和八腕幼体体长均表现为H组>C组>F组,体宽表现为无规律性;六腕幼体的胃长和胃宽均为F组>C组>H组,八腕幼体的胃长和胃宽均为C组>H组>F组。研究表明,一定近交水平 (F=0.25)的中间球海胆的受精、孵化,以及变态前幼体的体长、体宽均没有出现明显的近交衰退,但幼体胃长和胃宽的近交衰退现象比较明显。
关键词:中间球海胆;近交;幼体发育
近交是指有亲缘关系的个体间通过相互交配,使一个群体的纯合基因型增加的现象[1],近交可以纯化有害的隐性等位基因。不管是在自然条件下还是在人工养殖条件下,超过一定程度的近交通常会导致物种的某些表型性状值降低,这种表型性状值的降低称为近交衰退[2]。在水产动物方面,鱼[3-5]、 虾[6-7]、 贝类[8-9]和海胆[10]普遍存在着近交衰退现象。近交衰退在生物体的早期发育阶段表现最为明显,而到了发育后期,由于环境的选择效应淘汰了大部分衰退个体,会导致近交衰退的表现减弱很多[5,11-12]。
在水产领域,大量的引种实践表明,引种能够有效地调整养殖生态结构,增加经济效益和社会效益;但引种所带来的生态风险是毋庸回避的。近交和近交所引起的性状衰退便是其中之一。中间球海胆Strongylocentrotus intermedius具有重要的生态价值和经济价值,其性腺色泽好、味甜,在国际市场很受欢迎,目前已成为中国北方重要的水产养殖种类,出口价格高于其他经济海胆类[13-14]。自1989年大连水产学院从日本引进该种海胆以来,由于引种基础群体小、近亲繁殖和累代养殖,极有可能表现出生长缓慢、经济性状下降、抗逆性降低的近交衰退和种质下降。 迄今, 关于鱼[15]、 虾[16]、 贝类[17-20]近交的研究已有很多,但关于中间球海胆近交和近交衰退的研究尚未见报道。本研究中,作者研究了不同近交水平中间球海胆的受精、孵化和幼体发育阶段的差异,旨在探讨近交对中间球海胆繁殖和幼体生长发育的影响。
1.1 材料
试验用中间球海胆亲本选自大连海洋大学农业部北方海水增养殖学重点实验室2009年4月构建的F2代选育家系。试验用NIKON eclipse显微镜(型号50i)、NIKON相机 (型号YS100),用NISElements F和Adobe Photoshop软件处理图像。
1.2 方法
1.2.1 不同近交水平海胆的培育 2010年8月20日,以本实验室已经构建的F2代选育家系作为亲本,构建不同近交水平的海胆家系。用于构建选育家系基础群体的个体间均没有亲缘关系,并且在逐代选育过程中严格控制避免近交。本试验中利用上述F2代选育家系作为亲本,构建3组不同近交水平的海胆群体,即近交系数为0.25的全同胞近交组 (F),近交系数为 0.125的半同胞近交组(H),近交系数为0的对照组 (C),每组设置9个重复。
采用注射 KCl溶液法 (0.5 mol/L,1~2 mL/个)进行催产,分别收集每个亲胆的精液或卵液,严防混合。授精完成后,将各家系受精卵置于独立的70 L孵化槽内,经过2次洗卵去除多余的精子后进行孵化,孵化水温为18.0~19.0℃,光照度低于300 lx,不充气。受精后30 h左右至棱柱幼体,此时利用300目筛绢网捞取水面幼体,调整培育密度为2个/mL,水温与光照条件不变,微弱充气使幼体在水中分布均匀,投喂牟氏角毛藻,每天4次,投饵前镜检幼体的摄食消化情况,并以此确定投喂量,每天换水一次,换水量为1/2,每隔4天倒池一次。在以上条件下培育约21 d,幼体发育经二腕幼体、四腕幼体和八腕幼体期,各组的培育条件完全一致。
1.2.2 试验观察和取样测量 在海胆幼体发育到二细胞期、棱柱幼体、四腕、六腕和八腕5个时期时,分别用10 mL离心管对3个近交水平的每个重复进行随机取样,然后用体积分数为5%的福尔马林溶液杀死并固定幼体,每10 mL幼体样品加入1 mL固定液。固定后,将容器静置30 min,用塑料吸管吸取容器底部的幼体,滴至载玻片,将载玻片置于配备有数码相机、电脑和显微拍摄软件的显微镜下,在合适倍数物镜下调至清晰状态,物镜倍数以幼体的面积占视野面积的1/100~1/50为宜;打开图像软件,将幼体在屏幕中的图像调整至清晰,按照目镜倍数往图像中添加标尺,标尺实际长度Y,拍照并保存幼体照片。用Photoshop图像处理软件打开幼体照片,测量照片中标尺的相对长度X,随机选取幼体测量目标性状在照片中的尺寸Xi、目标性状的实际尺寸Yi,按下式换算:
Yi=Xi×Y/X。
在二细胞期和棱柱幼体期分别计算每个家系海胆的受精率 (各近交水平取样数大于800个)和孵化率 (各近交水平取样数大于800个);在四腕幼体期,测量畸形率 (各近交水平取样数大于 350个)、体长和体宽 (各近交水平取样数大于200个);在六腕和八腕幼体期,测量其体长、体宽、胃长、胃宽 (各近交水平取样数大于100个)(图1)。
受精率、孵化率和畸形率的计算公式如下:
受精率(%)=100×二细胞期总数/(二细胞期总数+未分裂的细胞数),
孵化率(%)=100×棱柱幼体总数/(棱柱幼体总数+未发育的细胞数),
畸形率(%)=100×畸形幼体总数/幼体总数。
图1 海胆幼体体长、体宽、胃长、胃宽的测量
Fig.1 M easurement of body length,body w idth, stomach length and stom ach w id th in sea urchin larvae
1.3 数据处理
用Excel软件对数据进行初步整理,以近交水平为影响因素,用SPSS 13.0软件对性状进行单因素方差分析,采用Duncan法对不同近交水平间的差异进行多重比较,显著性水平设为0.05。
2.1 不同近交水平海胆的受精率、孵化率和畸形率
3个近交水平海胆的受精率、孵化率和四腕幼体的畸形率见表1。从表1可见:3个近交水平海胆的受精率、孵化率和四腕幼体的畸形率均无显著性差异 (P>0.05);受精率和孵化率都在89%以上,而且变异系数都很小 (<8%),3个近交水平四腕幼体畸形率的变异系数均超过了20%。
2.2 不同近交水平海胆幼体的体长和体宽
从表2可见:四腕、六腕和八腕幼体的体长均表现为H组>C组>F组,其中,四腕幼体F组的体长显著小于C组和H组 (P<0.05),C组和H组间无显著性差异 (P>0.05);六腕幼体3组间的体长均无显著性差异(P>0.05),八腕幼体H组的体长显著大于C组和F组,C组和F组间无显著性差异(P>0.05)。不同近交水平四腕、六腕和八腕幼体的体宽表现为无规律性,其中,四腕幼体H组的体宽显著小于C组和F组 (P<0.05),C组和F组间无显著性差异 (P>0.05);六腕幼体F组的体宽显著大于C组和H组 (P<0.05),C组和H组间无显著性差异 (P>0.05);八腕幼体H组的体宽显著大于C组和F组 (P<0.05),C组和F组间无显著性差异 (P>0.05)。
表1 3个近交水平海胆的受精率、孵化率和畸形率
Tab.1 The fertility rate,hatching rate and aberration rate at the three inbreeding levels %
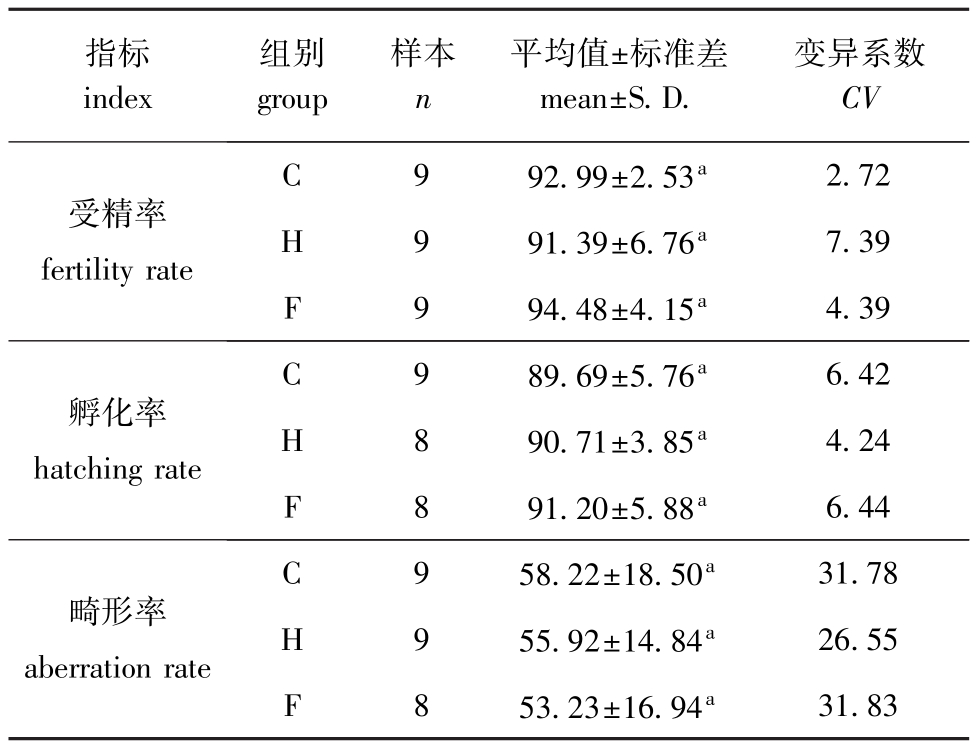
注:同列中标有不同小写字母者表示组间有显著性差异 (P<0.05),标有相同小写字母者表示组间无显著性差异 (P>0.05),下同。
Note:Themeanswith different letterswithin the same column are significant differences at the 0.05 probability level,and the means with the same letterswithin the same column are not significant differences,et sequentia.
指标index组别group样本n平均值±标准差mean±S.D.变异系数CV受精率fertility rate C 9 92.99±2.53a2.72 H 9 91.39±6.76a7.39 F 9 94.48±4.15a4.39孵化率hatching rate C 9 89.69±5.76a6.42 H 8 90.71±3.85a4.24 F 8 91.20±5.88a6.44畸形率aberration rate C 9 58.22±18.50a31.78 H 9 55.92±14.84a26.55 F 8 53.23±16.94a31.83
表2 3个近交水平四腕、六腕和八腕幼体的体长和体宽
Tab.2 The body length and body w id th of the echinopluterusw ith four,six and eight arms at the three inbreeding levels μm
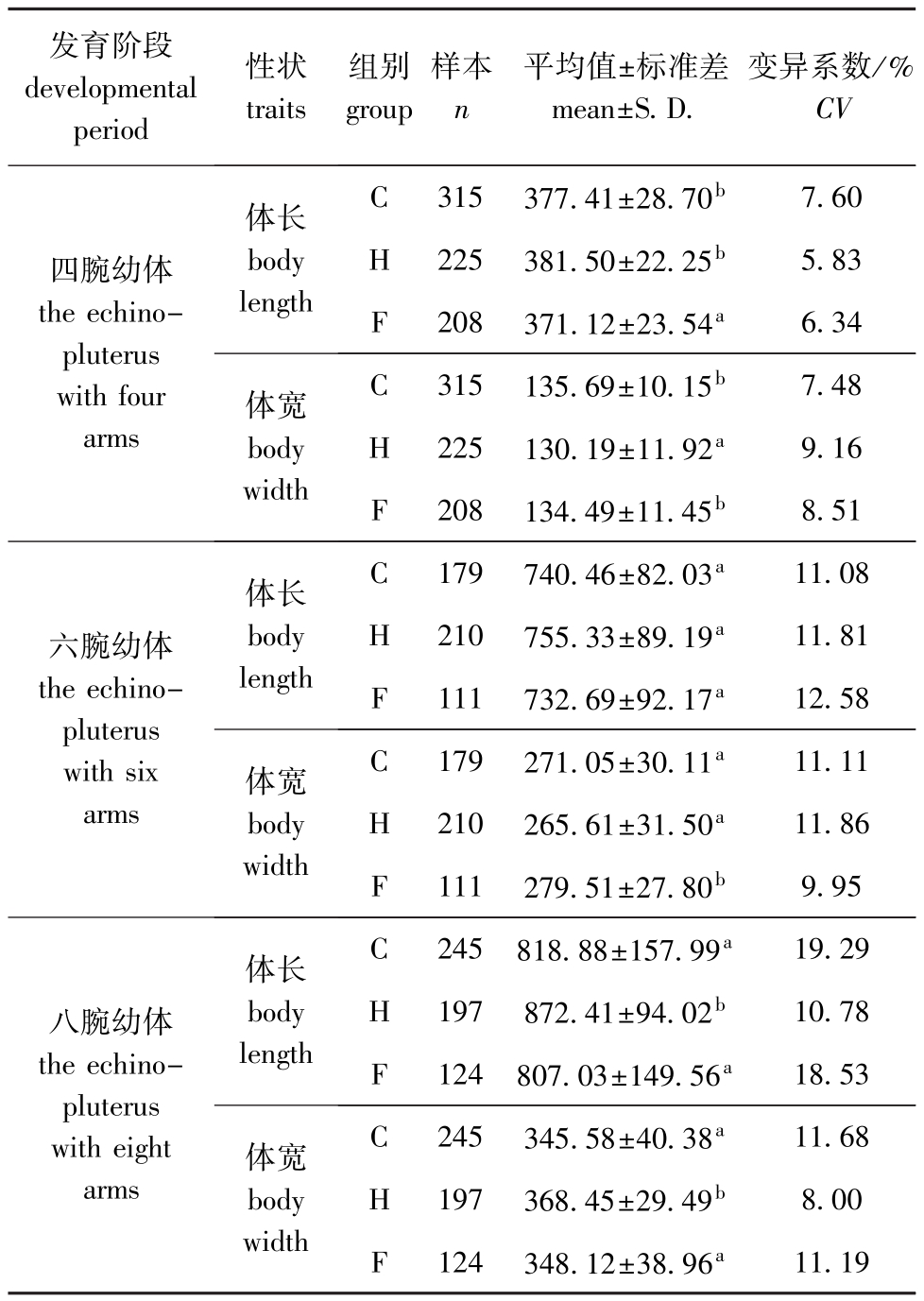
发育阶段developmental period性状traits组别group样本n平均值±标准差mean±S.D.变异系数/%CV四腕幼体the echinopluterus with four arms体长body length C 315 377.41±28.70b7.60 H 225 381.50±22.25b5.83 F 208 371.12±23.54a6.34体宽body width C 315 135.69±10.15b7.48 H 225 130.19±11.92a9.16 F 208 134.49±11.45b8.51六腕幼体the echinopluterus with six arms体长body length C 179 740.46±82.03a11.08 H 210 755.33±89.19a11.81 F 111 732.69±92.17a12.58体宽body width C 179 271.05±30.11a11.11 H 210 265.61±31.50a11.86 F 111 279.51±27.80b9.95八腕幼体the echinopluterus with eight arms体长body length C 245 818.88±157.99a19.29 H 197 872.41±94.02b10.78 F 124 807.03±149.56a18.53体宽body width C 245 345.58±40.38a11.68 H 197 368.45±29.49b8.00 F 124 348.12±38.96a11.19
2.3 不同近交水平海胆的胃长和胃宽
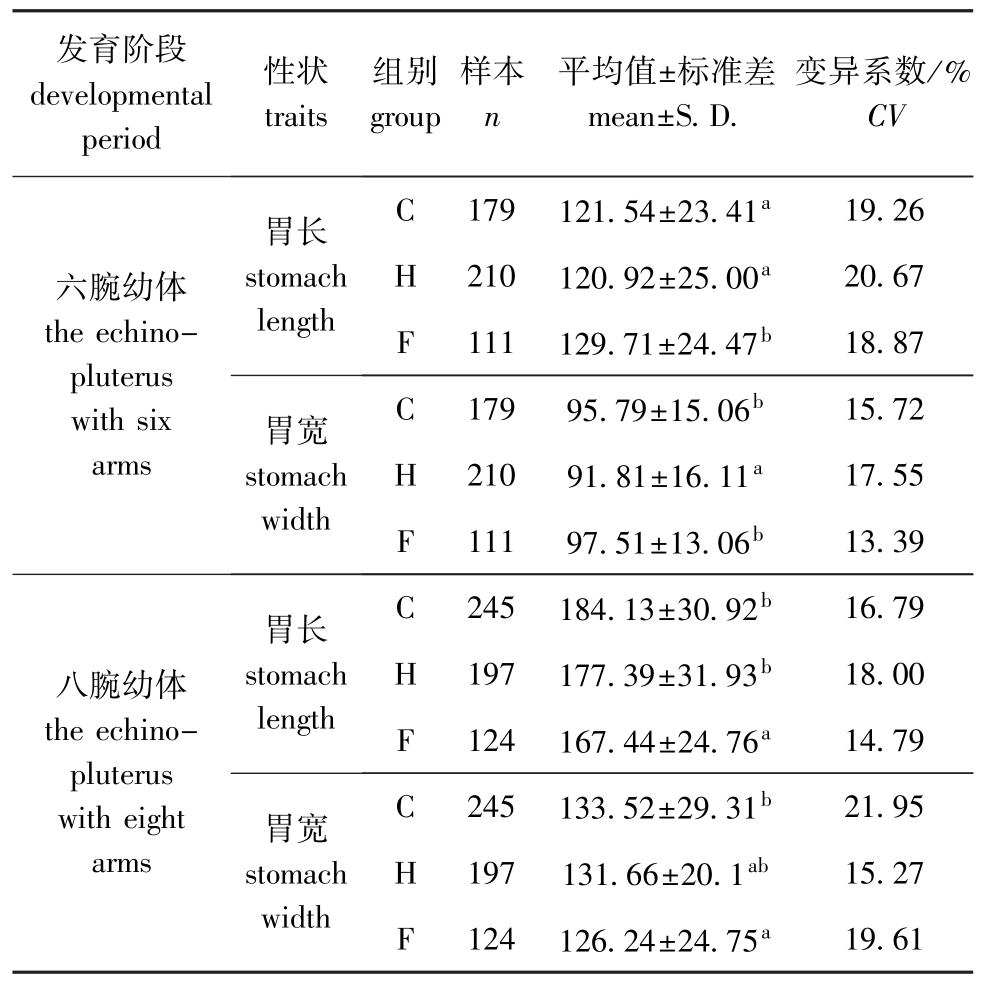
从表3可见:六腕幼体的胃长和胃宽均为F组>C组>H组,其中,F组的胃长显著大于C组和H组 (P<0.05),C组和 H组间无显著性差异(P>0.05),H组的胃宽显著小于C组和F组 (P<0.05),C组和F组间无显著性差异 (P>0.05);八腕幼体的胃长和胃宽均为C组>H组>F组,其中,F组的胃长显著小于C组和H组 (P<0.05), C组和H组间无显著性差异 (P>0.05),H组的胃宽与C组和F组无显著性差异(P>0.05),C组和F组间有显著性差异 (P<0.05)。
表3 3个近交水平六腕和八腕幼体的胃长和胃宽
Tab.3 The stomach length and stomach w idth of the echinopluterusw ith six and eight arms at the three inbreeding levels μm
?
虽然海洋生物的近交衰退现象被证实是普遍存在的,但不同物种近交衰退的时空表现不同[5,11,21]。因此,对不同海洋生物的相同或相似性状进行比较具有非常重要的研究价值。本研究结果表明,不同近交水平 (F=0、0.125、0.25)下中间球海胆的孵化率并没有显著差异。这一结果与对斑马鱼Brachydanio rerio[21]和虹鳟Salmo gairdneri[22]的研究结果相似。然而,与对这些海洋脊椎动物的研究结果不同的是,本试验中发现,不同近交水平海胆的受精率和四腕幼体畸形率也没有显著差异。Mrakovcic等[21]认为,近交能够显著地降低斑马鱼的受精率。Kincaid[22]的研究也发现,近交水平为0.25时能够显著增加虹鳟幼体的畸形率。作者认为,这可能与不同物种对近交所采取的应对策略不同有关。本研究结果表明,不同近交水平的中间球海胆受精率和孵化率的变异系数都很低(<8%),这表明中间球海胆受精率和孵化率在遗传上具有很强的保守性。不同近交水平海胆的受精率、孵化率和四腕幼体畸形率均没有显著差异很可能与这种保守性有关。
在海洋动物的生活史中,幼体发育对于生物的存活、生长和繁殖都是至关重要的。与鱼类近交的研究结果不同[5,21-22],本研究中发现,不同近交水平的海胆幼体各个发育时期的体长和体宽并没有出现明显的近交衰退现象。这与 Aulstad等[23-24]、Guo等[25]的研究结果一致。Aulstad等[23-24]研究发现,近交虽然导致虹鳟幼体的畸形率上升,但并不影响鱼苗正常生长,Guo等[25]的研究进一步证实了这一结论。他们在虹鳟幼体阶段 (256 d以前)没有观察到体质量的近交衰退现象。对于这一现象,一个合理的解释是虹鳟幼体存活率上出现的近交衰退抵消了幼体体质量的衰退,因此,在幼体阶段无法检测到近交衰退现象[12]。本试验中,虽然由于试验条件限制无法精确测量各近交水平幼体的死亡率,但观察到了不同近交水平的幼体在发育过程中的存活率有较大的差异。作者认为,不同近交水平下,不同死亡率对体长和体宽近交衰退的抵消,很可能是造成各近交水平海胆幼体体长和体宽没有显著差异的重要原因。当然,这还需进一步试验来验证。Anderson等[10]报道的近交紫球海胆Stronglyocentrotus purpuratus(F=0.25)幼体体长和体宽出现显著衰退现象,与本研究结果不一致。这表明不同物种 (即便同属)的近交响应策略也存在着较大差异。
虽然海胆变态的机制细节尚不清楚,但大量研究表明,其变态易受环境[26-27]特别是食物条件[28]的影响。Anger等[29]研究认为,在水生生物的幼体变态过程中要给与足够的饵料,否则将不能成功变态,最终将死去。胃是海胆幼体最主要的消化器官,Byrne等[30]的研究证实了胃对于海胆幼体变态具有重要的意义。本研究中发现,当海胆幼体发育至八腕时期,其胃表现出比较明显的近交衰退现象,近交系数为0.25的海胆胃长和胃宽要显著小于对照组。普遍认为,近交能够显著降低海洋无脊椎动物的变态率[31],然而其机制并不清楚。本研究中,作者推测近交组出现的胃衰退很可能是影响海胆变态成功率的重要原因。在海洋经济动物人工苗种培育生产实践中,变态率的高低直接决定着苗种的产量和产值。
参考文献:
[1] 马大勇,胡红浪,孔杰.近交及其对水产养殖的影响[J].水产学报,2006,29(6):849-856.
[2] Frankham R,Gilligan D M,Morris D,et al.Inbreeding and extinction:effects of purging[J].Conserv Genetics,2001,2(3):279-284.
[3] Arkush K D,Giese A R,Mendonca H L,et al.Resistance to three pathogens in the endangered winter-run chinook salmon(Oncorhynchus tshawytscha):effects of inbreeding and major histocompatibility complex genotypes[J].Canadian Journal of Fisheries and Aquatic Sciences,2002,59(6):966-975.
[4] Gallardo J A,Garcia X,Lhorente J P,et al.Inbreeding and inbreeding depression of female reproductive traits in two populations of coho salmon selected using BLUP predictors of breeding values[J].Aquaculture,2004,234(1/4):111-122.
[5] Fessehaye Y,Komen H,Rezk M A,et al.Effects of inbreeding on survival,body weight and fluctuating asymmetry(FA)in Nile tilapia,Oreochromisniloticus[J].Aquaculture,2007,264(1/4):27-35.
[6] 孔杰,金武,栾生,等.水产动物选择育种的近交分析[J].自然科学进展,2009,19(9):917-923.
[7] Keys S J,Crocos P J,Burridge C Y,et al.Comparative growth and survival of inbred and outbredPenaeus(Marsupenaeus)japonicus, reared under controlled environment conditions:indications of inbreeding depression[J].Aquaculture,2004,241(1/4):151-168.
[8] Langdon C,Evans F,Jacobson D,et al.Yields of cultured Pacific oystersCrassostrea gigasThunberg improved after one generation of selection[J].Aquaculture,2003,220(1/4):227-244.
[9] Lind C E,Evans B S,Knauer J,et al.Decreased genetic diversity and a reduced effective population size in cultured silver-lipped pearl oysters(Pinctada maxima)[J].Aquaculture,2009,286(1/ 2):12-19.
[10] Anderson D,Hedgecock D.Inbreeding depression and growth heterosis in larvae of the purple sea urchinStronglyocentrotus purpuratus(Stimpson)[J].Journal of ExperimentalMarine Biology and Ecology,2010,384:68-75.
[11] Lande R,Schemske DW.The evolution of self-fertilization and inbreeding depression in plants:a population genetic model[J].Evolution,1985,39(1):24-40.
[12] Gjerde B,Gunnes K,Gjedrem T.Effect of inbreeding on survival and growth in rainbow trout[J].Aquaculture,1983,34(3/4): 327-332.
[13] 秦艳杰,李霞,吴立新,等.饥饿和再投喂对中间球海胆代谢和生长的影响[J].大连海洋大学学报,2011,26(6):521-525.
[14] 苏延明,项福建,宋坚,等.虾夷马粪海胆反季节繁育技术的研究[J].大连水产学院学报,2008,23(2):136-140.
[15] 王磊,陈松林,田永胜,等.牙鲆回交、全同胞近交及其亲本家系的微卫星研究[J].渔业科学进展,2011,32(3):38-43.
[16] 张洪玉,罗坤,孔杰,等.近交对中国明对虾生长、存活及抗逆性的影响[J].中国水产科学,2009,16(5):744-750.
[17] 刘振,闫喜武,霍忠明,等.蛤仔白斑马品系的家系选择及近交效应研究[J].大连海洋大学学报,2013,28(4):355-360.
[18] 王宇,李莉,张守都,等.海湾扇贝杂交、近交和自交家系的微卫星标记偏分离分析[J].海洋科学,2012,36(8):109-115.
[19] 闫喜武,孙欣,张跃环,等.菲律宾蛤仔奶牛蛤品系两个世代的杂交与近交效应[J].水产学报,2011,35(5):682-691.
[20] 高悦勉,李国喜,赵银丽.大连沿海虾夷扇贝养殖群体遗传结构的研究[J].大连水产学院学报,2004,19(2):142-145.
[21] Mrakovcic M,Haley L E.Inbreeding depression in the zebra fishBrachydanio rerio(Hamilton:Buchanan)[J].Journal of Fish Biology,1979,15(3):323-327.
[22] Kincaid H L.Inbreeding in rainbow trout(Salmo gairdneri)[J]. Journal of the Fisheries Research Board of Canada,1976,33 (11):2420-2426.
[23] Aulstad D,Gjedrem T,Skjervold H.Genetic and environmental sources of variation in length and weight of rainbow trout(Salmo gairdneri)[J].Journal of the Fisheries Research Board of Canada,1972,29(3):237-241.
[24] Aulstad D,Kittelsen A.Abnormal body curvatures of rainbow trout (Salmo gairdneri)inbred fry[J].Journal of the Fisheries Research Board of Canada,1971,28(12):1918-1920.
[25] Guo Shengsu,Liljedahl L E,Gall G A E.Effects of inbreeding on growth and reproductive traits in rainbow trout(Oncorhynchus mykiss)[J].Aquaculture,1996,142(3/4):139-148.
[26] Diaz-Perez L,Carpizo-Ituarte E.Effect of thermal stress on survival and delay ofmetamorphosis in larvae of the purple sea urchinStrongylocentrotus purpuratus[J].Ciencias Marinas,2011, 37(4):403-414.
[27] Salas-Garza A,Pares-Sierra G,Ponce-Marquez M A.Culture of benthic diatoms and effect of dissolved organic carbon on metamorphosis of the“white sea urchinLytechinus pictus(Verrill)”[J].Aquaculture Research,2010,41(4):535-544.
[28] Xing Ronglian,Wang Changhai,Cao Xuebin,et al.The potential value of different species of benthic diatoms as food for newly metamorphosed sea urchinStrongylocentrotus intermedius[J]. Aquaculture,2007,263(1/4):142-149.
[29] Anger K,Dawirs R R,Anger V,et al.Effects of early starvation periods on zoeal development of brachyuran crabs[J].The Biological Bulletin,1981,161(2):199-212.
[30] Byrne M,Sewell M A,Prowse T A A.Nutritional ecology of sea urchin larvae:influence of endogenous and exogenous nutrition on echinopluteal growth and phenotypic plasticity inTripneustes gratilla[J].Functional Ecology,2008,22(4):643-648.
[31] Longwell A C,Stiles S S.Gamete cross incompatibility and inbreeding in the commercial American oysterCrassostrea virginicaGmelin[J].Cytologia,1973,38:521-533.
Effect of inbreeding on fertility rate,hatchability and larva development of sea urchin Strongylocentrotus intermedius
Abstract:In the present study,we investigated the effects of inbreeding on fertility rate,hatchability and larva development of sea urchin Strongylocentrotus intermedius by establishing three inbreeding level(F=0,0.125,0.25) groups(C,H and F).The results showed that the influence of inbreeding on fertility rate,hatchability and aberration rate of four arms larvawere not significant(P>0.05).The order of body length of larvae with four-arms,sixarms and eight-arms at the different inbreeding levels was ranged as H group>C group>F group.The change in body width has no remarkable regularity.The order of length and width of stomach in six arms larvae was ranged as F group>C group>H group.The length and width of stomach in eight arms larvae was expressed as C group>H group>F group.The results showed that the inbreeding level of F=0.25 was not harmful to the fertility rate,hatchability and larva development of the sea urchinStrongylocentrotus intermediusexcept the development of stomach.
Keywords:Strongylocentrotus intermedius;inbreeding;larva development
中图分类号:S968.22
文献标志码:A
文章编号:2095-1388(2013)05-0445-05
收稿日期:2013-01-31
基金项目:国家 “863”计划项目 (2012AA10A412)