柱状黄杆菌对草鱼TLRs基因表达水平的影响
隗黎丽,吴华东,熊六凤
(江西农业大学动物科学技术学院,江西南昌330045)
摘要:用柱状黄杆菌Flavobacterium columnareG4株感染草鱼Ctenopharyngodon idella,分别在感染1、4、7 d后提取感染组和对照组草鱼鳃、脾脏、肝脏、肠道和头肾5种组织的总RNA,并采用半定量RT-PCR方法检测不同组织中Toll样受体3(TLR3)、Toll样受体7(TLR7)和Toll样受体22(TLR22)3个抗病毒免疫基因的表达变化。结果表明:注射柱状黄杆菌7 d后,TLR3在草鱼鳃、肝脏、肠道和头肾4种组织中的表达显著上调 (P<0.05);注射柱状黄杆菌4 d和7 d后,TLR7和TLR22在5种组织中的表达均显著上调(P<0.05);而注射柱状黄杆菌1 d后,TLR22在鳃、脾脏和肝脏组织中的相对表达量就显著上调 (P<0.05)。研究表明,TLR3、TLR7和TLR22基因在机体应对细菌感染的免疫反应过程中发挥着重要的作用。
关键词:草鱼;柱状黄杆菌;Toll样受体基因;基因表达
草鱼Ctenopharyngodon idella是中国重要的淡水鱼类养殖品种之一,具有生长快、对饲料蛋白需求低、肉质好等优点。随着养殖密度的增大和养殖环境的恶化,草鱼细菌性烂鳃病频繁暴发,造成巨大的经济损失。烂鳃病的病原菌为柱状黄杆菌Flavobacterium columnare,主要危害鱼苗和鱼种,死亡率非常高,尤以草鱼为甚[1]。免疫防治是当前鱼类病害最理想的控制手段[2],因此,对鱼类免疫相关的基础研究显得尤为重要。鱼类在受病菌侵染时,自身通过一系列信号传递启动抗病防御系统,有序地引发一系列免疫系统的变化,产生抵抗作用,这一切都源于免疫基因的转录调控,因此,研究免疫相关基因在转录水平的表达,有助于认识抗病的分子机理。研究发现,Toll样受体 (Toll-like receptors,TLRs)作为一类模式识别受体 (pattern recognizing receptor,PRR),在免疫应答和炎性反应中发挥着重要作用[3]。TLR可通过识别外源性微生物,启动先天性免疫反应,清除侵入的病原微生物[4],其最突出的生物学功能是促进细胞因子的合成与释放,从而引发炎症反应 (inflammation)[5]。Toll样受体首次在果蝇体内发现并被报道以来,在哺乳动物中已经发现了至少13种TLR家族成员,在水产动物中已发现了几十种TLR家族成员,其中一些为鱼类所特有的TLR家族成员[6]。本研究中,作者采用半定量RT-PCR方法,对柱状黄杆菌引起的草鱼Toll样受体3(TLR3)、Toll样受体7(TLR7)和Toll样受体22(TLR22)基因在不同组织中的表达情况进行检测分析,旨在探索草鱼对柱状黄杆菌的免疫反应规律。
1 材料与方法
1.1 材料
柱状黄杆菌G4株由中国科学院水生生物研究所鱼类免疫学与寄生虫实验室馈赠,该菌的培养条件及培养基配方参考文献 [7]中的方法。
试验用草鱼于2011年1月购买于江西省新建县一渔场,体质量为15~20 g。将购买的草鱼在实验室内暂养,每日投喂一次商品颗粒饲料,水温保持在25℃左右。选取体表完整、健康的暂养草鱼,分别置于玻璃缸 (100 cm×60 cm×50 cm)中饲养。
将草鱼分为试验组和对照组,试验组鱼经胸腔注射柱状黄杆菌G4株 (1.0×107cfu/mL)100 μL,对照组鱼注射灭菌的PBS(0.1 mol/L,pH 7.2)。经诱导1、4、7 d后取样,每个时间点各取4条鱼,分别取鳃、头肾、脾脏、肝脏和肠道组织,用于提取RNA。
1.2 方法
1.2.1 RNA的提取 用Trizol(Invitrogen)提取各组织的总RNA。各组织样品经液氮冷冻后加入1 mL的Trizol,再用匀浆器进行匀浆,匀碎后,倒入无RNase的离心管中,室温下静置10 min,离心(12 000 g,10 min,4℃);取上清,加入200 μL的氯仿,剧烈摇动15 s,室温静置3 min,离心(12 000 g,15 min);再取上清,转移到干净的离心管中,加入500 μL的异丙醇,混匀后,室温静置10 min,离心 (10 000 g,10 min,4℃);去掉上清,用体积分数为75%的乙醇洗涤沉淀,离心(7 500 g,5 min,4℃);最后加入体积分数为100%的乙醇,于-80℃下保存备用。用12 g/L TAE琼脂糖凝胶电泳检测总RNA的质量。
1.2.2 cDNA第一链的合成 取制备的RNA样品,用20 μL DEPC水溶解,用紫外分光光度计测定每个RNA样品的浓度。取5 μg总RNA进行反转录。25 μL反转录体系中加入5 μg总RNA模板,再加入1 μL Oligo(dT)18,于 70℃下反应5 min,冰上冷却,离心;然后加入 5×Reaction Buffer 4 μL,Ribolock TM Ribonuclease Inhibitor 1 μL,10 mmol/L dNTP mix 2 μL,RevertAidTMMMuLV Reverse Transcriptase 1 μL;最后再加入DEPC水使反应体积达到25 μL。于42℃下反应60 min,反应结束后将产物分装,于-20℃下冷冻保存备用。
1.2.3 用RT-PCR法检测TLRs mRNA的相对表达 本试验中草鱼TLR3、TLR7和TLR22基因的引物设计分别参考文献 [8-10]中的方法,详见表1。以草鱼cDNA为模板,用所设计的引物进行PCR扩增,然后进行琼脂糖凝胶电泳检测。20 μL反应体系中包含1.0 μL cDNA模板,正反向PCR引物各0.5 μL(10 μmol/L),0.5 μL dNTP(10 μmol/L),2.5 μL 10 ×Buffer,0.2 μL ExTaqE (TaKaRa公司产品),加灭菌水补充至20 μL。反应程序如下:94℃下预变性5 min;94℃下变性30 s,58℃下退火30 s,72℃下延伸40 s,共进行25~45个循环;最后在72℃下延伸5 min,为避免扩增达到平台期,分别用琼脂糖凝胶电泳检测25、30、35、40个循环时的扩增产物,最后选择最佳的循环数。本试验中PCR产物用EB染色[11],用12 g/L TAE琼脂糖凝胶进行电泳,并用凝胶电泳显像仪拍照。
表1 目的基因及内参基因的引物
Tab.1 Primers of target and internal control genes

基因gene登录号GenBank No.正向引物(5'-3') forward primer(5'-3')反向引物(5'-3') reverse primer(5'-3')片段大小/bp fragment size Toll-like receptor 3 DQ864497 F:ACCAGGTAATGGAGAACAATCG R:GCCAGTAGAGAACACAGCGAG 129 Toll-like receptor 7 FJ610253 F:GAGCATACAGTTGAGTAAACGCAC R:TCTCCAAGAATATCAGGACGATAA 143 Toll-like receptor 22 FJ547498 F:TCGTTTGTCTGGCATTTTCTG R:CAAGGCTCGTCTTCGGTGT 146 β-actin M25013 F:CCTTCTTGGGTAGGAGTCTTG R:AGAGTATTTACGCTCAGGTGGG 221
1.3 数据处理
用凝胶图像分析软件Quantity One 4.62对所获得的TLRs 3个基因及β-actin进行光密度分析。各目的基因mRNA的相对表达量以目的基因光密度值与内参基因β-actin光密度的比值表示。试验数值均用平均值±标准差表示,采用SPSS 11.5进行单因素方差分析,用t检验法进行组间多重比较,以P<0.05表示有显著性差异。
2 结果
2.1 TLR3 mRNA在草鱼各组织中的表达变化
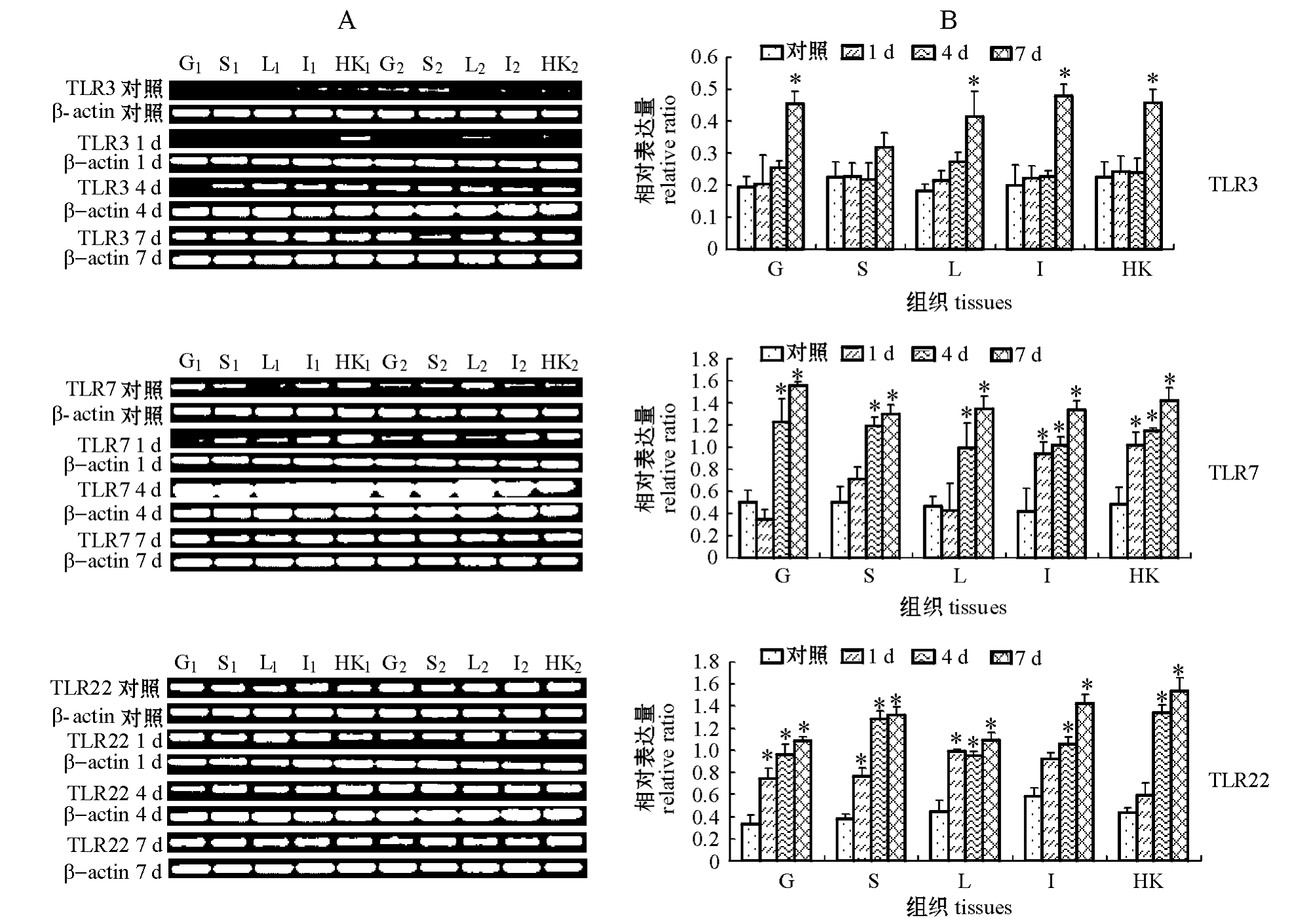
从图1可见:注射后各时间点,TLR3基因在对照组和试验组草鱼各组织中均能表达,注射柱状黄杆菌1 d和4 d后,TLR3基因在草鱼鳃、脾脏、肝脏、肠道和头肾5种组织中的表达变化不明显,但注射7 d后,TLR3基因在5种组织中的表达均呈现上调的趋势;除脾脏外,TLR3基因在其他4种组织中的表达均显著上调 (P<0.05),其中TLR3基因在肠道中的表达变化最大,比对照组升高了2.38倍。
2.2 TLR7 mRNA在草鱼各组织中的表达变化
从图1可见:注射后各时间点,TLR7在对照组和试验组草鱼各组织中均能表达,注射柱状黄杆菌4 d和7 d后,TLR7在5种组织中的表达均较对照组显著上调 (P<0.05),其中,TLR7在鳃组织中的表达变化最大,比对照组升高了3.09倍;注射柱状黄杆菌1 d后,TLR7在肠道和头肾组织中的也呈现显著上升的趋势 (P<0.05)。
2.3 TLR22 mRNA在草鱼各组织中的表达变化
从图1可见:注射后各时间点,TLR22在对照组和试验组草鱼各组织中也均能表达,注射柱状黄杆菌不同时间后,TLR22在5种组织中的表达均呈现上升的趋势,尤其是在注射柱状黄杆菌4 d和7 d后,TLR22在5种组织中的表达均较对照组显著上调 (P<0.05),而注射柱状黄杆菌 1 d后, TLR22在鳃、脾脏和肝脏组织中的表达就较对照组显著上调 (P<0.05)。
3 讨论
TLRs是天然免疫在进化过程中产生的天然免疫识别分子,能够识别病原相关分子模式 (pathogen-associated molecular patterns,PAMPs)[12]。Toll蛋白最早在果蝇体内被发现,它是果蝇在胚胎发育过程中调节果蝇背腹部体轴必需的成分蛋白[13]。 20世纪90年代中期,人们又发现该蛋白能感受到入侵的病原体,并且在此基础上使果蝇分泌多种抗微生物感染的多肽清除病原体,Toll受体由此得名[14]。活化的TLRs能激活T细胞,启动获得性免疫反应[15]。因此,TLRs在先天性免疫和获得性免疫两种机制中均起着重要作用。
研究表明,TLR3基因是双链RNA病毒模式识别受体之一,在识别双链RNA病毒的过程中起着重要作用[16]。Su等[8]在注射草鱼出血病病毒(GCRV)1、2、3、4、5、6、7 d后都能检测到脾脏中的TLR3基因显著诱导表达。在斑马鱼和虹鳟中,病毒也均能诱导 TLR3基因的表达[16-17]。TLR3除了参与抗病毒反应外,还能起到抵御细菌侵袭的作用。Bilodeau等[18]发现,当斑点叉尾鮰暴露于有毒力的爱德华氏菌Edwardsiella ictaluri液中时,鱼体内的TLR3表达被激活。本研究中发现,用柱状黄杆菌攻毒草鱼7 d后,TLR3在5种组织中的表达均能显著被诱导激活。高天珩等[19]发现,黄芪皂甙可以显著提高TLR3基因在斑点叉尾鮰肝脏中的表达量,且呈现较为明显的剂量效应,这些研究充分表明了TLR3是鱼类先天免疫系统中重要的组成部分,不仅在识别病毒的过程中起着重要作用,而且在抵御病原菌的过程中可被激活。
TLR7作为TLRs家族的一员,在病毒ssRNA的识别以及感染后的信号传导过程中起着重要作用[20]。本研究中发现,草鱼在感染柱状黄杆菌4 d和7 d后,TLR7在鳃、脾脏、肝脏、肠道和头肾5种组织中的表达均显著上调,这与张荣芳[21]的研究结果比较一致。张荣芳[21]在用草鱼呼肠孤病毒感染草鱼后,TLR7在草鱼鳃、肠、肝脏、脾脏和中肾等组织中的表达均上调。这种在细菌和病毒感染后表达被激活的现象显示了TLR7在先天性免疫反应中应答对抗细菌和病毒的免疫效应。
TLR22是鱼类所特有的TLR识别受体之一,它是定位于细胞膜上识别长双链RNA的一个识别受体,功能和哺乳动物细胞膜上TLR3类似[22-23]。鱼类TLR22可以识别双链RNA和细菌衍生物,用poly(I∶C)刺激诱导大黄鱼后,其头肾、脾脏组织以及头肾细胞中的TLR22基因的表达也显著上调[24]。在注射草鱼出血病病毒 (GCRV)2 d后,草鱼头肾和脾脏中TLR22的表达显著上调[25]。热灭活的杀鲑气单胞菌以及在培养的巨噬细胞中活的龟分支杆菌均能显著上调TLR22基因的表达[26]。在体外培养的外周血淋巴细胞、脾脏和头肾中,灭活的杀鲑气单胞菌可诱导TLR22基因表达上升8倍[27]。本研究中也发现,在用柱状黄杆菌感染草鱼后,5种不同免疫组织中TLR22的表达均上调,尤其是在感染4 d和7 d后,TLR22相对表达量显著上调。这些研究均表明了TLR22基因在免疫反应过程中起着重要作用。
柱状黄杆菌是引起草鱼流行性烂鳃病的一种重要病原,也是引起草鱼大规模死亡的一个重要病因。在以往的研究中,已知TLR3、TLR7和TLR22在抗病毒的过程中起着非常重要的作用,而从本研究结果来看,这些TLRs基因在鱼类抵御细菌的过程中同样也发挥着积极的作用,TLRs家族及其信号通路可能参与了机体对柱状黄杆菌的识别和防御,但具体的机制还有待进一步研究。
致谢:在样品准备过程中,中国科学院水生生物研究所鱼类免疫学与寄生虫学实验室馈赠了柱状黄杆菌,同时,李万雄等同学在养鱼和攻毒试验中提供了帮助,在此一并表示感谢!
参考文献:
[1] 谢海侠.柱状黄杆菌毒力相关因子的研究[D].武汉:中国科学院水生生物研究所,2005.
[2] Gill R W,Sanseau P.Rapid in silico cloning of genes using expressed sequence tags(ESTs)[J].Biotechnology Annual Review, 2000,5:25-44.
[3] Herbst-Kralovetz M M,Pyles R B.Quantification of poly(I:C) mediated protection against genital herpes simplex virus type 2 infection[J].Journal of Virology,2006,80(20):9988-9997.
[4] Akira S,Uematsu S,Takeuchi O.Pathogen recognition and innate immunity[J].Cell,2006,124:783-801.
[5] Foster S,Hargreaves D,Medzhitov R.Gene-specific control of inflammation by TLR-induced chromatin modifications[J].Nature, 2007,447:972-978.
[6] Yniv P.Toll-like receptors in bony fish:from genomics to function [J].Developmental and Comparative Immunology,2011,35: 1263-1272.
[7] 王良发,谢海侠,张金,等.我国淡水鱼类柱形病原菌柱状黄杆菌的遗传多样性[J].水生生物学报,2010,34(2):367-377.
[8] Su J,Jang S,Yang C,et al.Genomic organization and expression analysis of Toll-like receptor 3 in grass carp Ctenopharyngodon idella[J].Fish and Shellfish Immunology,2009,27:433-439.
[9] Yang C,Su J,Zhang R,et al.Identification and expression profiles of grass carp Ctenopharyngodon idella TLR7 in responses to double-stranded RNA and virus infection[J].Journal of Fish Biology,2012,80:2605-2622.
[10] Su J,Heng J,Huang T,et al.Identification,mRNA expression and genomic structure of TLR22 and its association with GCRV susceptibility/resistance in grass carp Ctenopharyngodon idella [J].Developmental and Comparative Immunology,2012,36:450-462.
[11] 赵文,毕进红,韩婷婷,等.西藏拟溞遗传多样性的初步研究[J].大连海洋大学学报,2011,26(1):108-113.
[12] Subramaniam S,Stansberg C,Cunningham C.The interleukin 1 receptor family[J].Developmental and Comparative Immunology, 2004,28:415-428.
[13] Anderson K V,Jurgens G,Nusslein-Volhard C.Establishment of dorsal-ventral polarity in the Drosophila embryo:genetic studies onthe role of the Toll gene product[J].Cell,1985,42:779-789.
[14] Lemaitre B,Nicolas E,Michaut L,et al.The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults[J].Procedings of the National Academy of Science,1996,86:973-983.
[15] Lieu Y K,Kumar A,Pajerowski A G,et al.Requirement of c-myb in T cell development and in mature T cell function[J].Proc Natl Acad Sci USA,2004,101(41):14853-14858.
[16] Phelan P E,Mellon M T,Kim C H.Functional characterization of full-length TLR3,IRAK-4,and TRAF6 in zebrafish Danio rerio [J].Molecular Immunology,2005,42:1057-1071.
[17] Rodriguez M F,Wiens G D,Purcell M K et al.Characterization of Toll-like receptor 3 gene in rainbow trout Oncorhynchus mykiss [J].Immunogenetics,2005,57:510-519.
[18] Bilodeau A L,Waldbieser G C.Activation of TLR3 and TLR5 in channel catfish exposed to virulent Edwardsiella ictaluri[J].Developmental and Comparative Immunology,2005,29:713-721.
[19] 高天珩,蒋广霞,许志强,等.黄芪皂甙对斑点叉尾鮰非特异性免疫和抗病力的影响[J].大连海洋大学学报,2010,25(6): 560-564.
[20] Diebold S S,Kaisho T,Hemmi H,et al.Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA [J].Science,2004,303:1529-1531.
[21] 张荣芳.草鱼TLR7基因的克隆与病毒感染条件下的表达研究[D].杨凌:西北农林科技大学,2011.
[22] Roach J C,Glusman G,Rowen L,et al.The evolution of vertebrate Toll-like receptors[J].Procedings of the National Academy of Science,2005,102(27):9577-9582.
[23] Matsuo A,Oshiumi H,Tsujita T,et al.Teleost TLR22 recognizes RNA duplex to induce IFN and protect cells from birnaviruses[J]. Immunology,2008,181:3474-3485.
[24] Xiao X Q,Qin Q W,Chen X H.Molecular characterization of a Toll-like receptor 22 homologue in large yellow croaker Pseudosciaena crocea and promoter activity analysis of its 5'-flanking sequence[J].Fish and Shellfish Immunology,2011,30:224-233.
[25] Lv J,Huang R,Li H,et al.Cloning and characterization of the grass carp Ctenopharyngodon idella Toll-like receptor 22 gene,a fish-specific gene[J].Fish and Shellfish Immunology,2012,32: 1021-1031.
[26] Stafford J L,Ellestad K K,Magor K E,et al.A Toll-like receptor (TLR)gene that is up-regulated in activated goldfish macrophages[J].Developmental and Comparative Immunology,2003,27: 685-698.
[27] Rebl A,Siegl E,Küllner B,et al.Characterization of twin tolllike receptors from rainbow trout Oncorhynchus mykiss:evolutionary relationship and induced expression by Aeromonas salmonicida [J].Developmental and Comparative Immunology,2007,31:499-510.
Effects of bacterial pathogen Flavobacterium columnare on expression of Toll receptors genes in grass carp Ctenopharyngodon idella
WEI Li-li,WU Hua-dong,XIONG Liu-feng
(College of Animal Science and Technology,Jiangxi Agricultural University,Nanchang 330045,China)
Abstract:In the present study,the expressions of three Toll receptor genes TLR3,TLR7 and TLR22 were investigated in gill,spleen,hepatopancreas,intestine and head kidney in grass carp Ctenopharyngodon idella intraperitoneally injected with bacterial pathogen Flavobacterium columnare G41,4 and 7 days after injection by semi-quantified RT-PCR which were analyzed by electrophoresis on 12 g/L TAE agarose gels.The densitometry was used by Quantity One 4.62,and in the control group,the fish were injected intraperitoneally with sterile phosphate buffer solution(PBS).The results showed that the expression of TLR3 gene was found to be significantly increased in the gill,hepatopancreas,intestine and head kidney 7 days after injection compared with the fish in the control group, and the expressions of TLR7 and TLR22 genes were significantly upregulated in 5 different tissues 4 and 7 days after injection.The expression of TLR22 gene was increased significantly in gill and spleen and hepatopancreas compared with the control fish only 1 day after injection,indicating that TLR3,TLR7 and TLR22 gens are effectively activated after administration by the pathogen F.columnare.
Key words:Ctenopharyngodon idella;Flavobacterium columnare;TLRs;gene expression
中图分类号:S185
文献标志码::A
文章编号:2095-1388(2013)04-0378-05
收稿日期:2012-12-23
基金项目:江西省教育厅科技计划项目 (GJJ11087);江西省科技计划项目 (20114BAB214002)
作者简介:隗黎丽 (1976-),女,讲师,博士。E-mail:hbliliwei@163.com
通信作者:熊六凤 (1970-),男,副教授,博士。E-mail:xlf227@126.com