
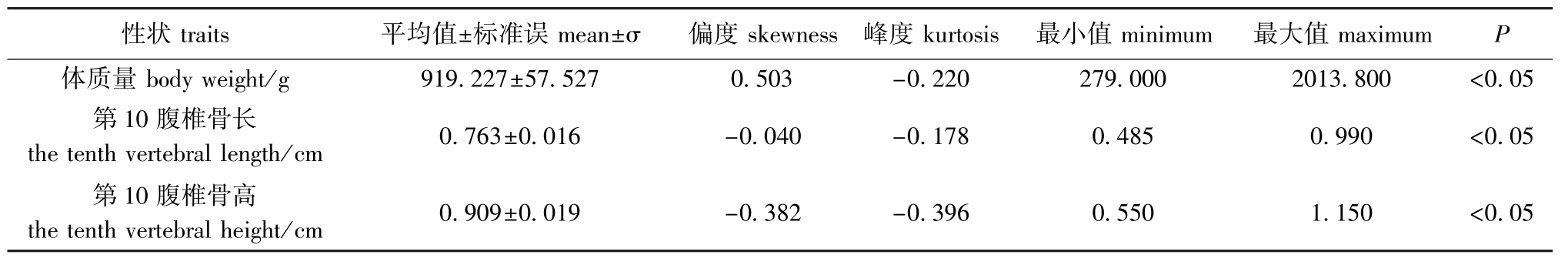
图1 牙鲆X-射线图
Fig.1 The X-ray photography of Japanese flounder Paralichthys olivaceus
摘要:利用94对微卫星标记,对52尾双单倍体牙鲆Paralichthys olivaceus第10腹椎骨的长度和高度与微卫星标记的相关性进行分析。结果表明:微卫星标记Poli1222TUF、Poli1418TUF、Po1、Po91、Poli1498TUF、Poli11795TUF与第10腹椎骨长显著相关 (P<0.05);Poli1406TUF与第10腹椎骨高显著相关 (P<0.05); Poli1143TUF、Poli1356TUF和Poli1388TUF则与第10腹椎骨长和骨高均显著或极显著相关 (P<0.05或P<0.01)。研究表明,这些标记可以作为牙鲆辅助育种的参考。
关键词:牙鲆;雌核发育;双单倍体;微卫星标记;腹椎骨
牙鲆Paralichthys olivaceus俗称牙片、偏口、比目鱼,是分布于中国沿海的大型经济鱼类,其个体硕大、生长快速、肉质细嫩鲜美,具有很高的经济价值。由于近年来的过度捕捞和环境污染,牙鲆自然资源大幅度下降,为了满足市场需求,人们开始了大规模的人工养殖牙鲆。随着累代养殖,近亲繁殖和性状衰退现象明显,因此,选育优良性状的牙鲆,对其养殖业的发展具有重要意义[1]。
人工诱导雌核发育是快速建立纯系的有效手段。通过抑制第一次卵裂可以获得有丝分裂雌核发育二倍体,即双单倍体 (double haploid,DH)。同一母本的DH子代都是纯合体,但后代个体之间基因型互不相同,是进行遗传连锁分析的好材料[2-3]。微卫星又称为短串联重复序列 (simple tandem repeats,STR)或简单重复序列 (simple sequence repeat,SSR),具有串联重复单元的重复次数在个体间变异性高、数量丰富、稳定性好、多态性高、呈共显性遗传和遵循孟德尔分离定律等特点[4],微卫星标记作为分析重要经济性状遗传连锁关系的理想标记,现已广泛应用于水产动物的遗传图谱构建[5-6]、遗传多样性分析[7-8]、群体遗传结构分析[9-10]、 基因定位[11-12]、 QTL分析[13]等。
骨骼的生长受遗传因素的影响较大,受外环境影响较小[14-15]。所以寻找骨骼参数与经济性状紧密相关的标记将会更准确、更稳定。牙鲆体长主要由头骨、脊椎骨和尾骨组成,体高由脊椎骨高和肋骨长组成。牙鲆的脊椎骨由腹椎骨和躯椎骨组成,第10腹椎骨为腹椎骨的最后一块,也是所有脊椎骨中较大的一块,因此是脊椎骨中的重要骨骼。本研究中,作者以DH牙鲆第10腹椎骨的骨长和骨高为研究对象,分析微卫星位点与其相关性,筛选出与这些性状相关的分子标记,旨在为牙鲆经济性状的QTL定位及分子标记辅助育种提供基础数据。
1.1 材料
1.1.1 试验鱼 采用中国水产科学研究院北戴河中心试验站选育的野生牙鲆亲鱼为试验材料。采用王凯等[16]诱导雌核发育的方法,用紫外线灭活的真鲷Pagrosomus major精子与牙鲆雌鱼的同一批卵混合,静置3 min后,加入17℃的海水激活牙鲆雌鱼卵,1 h后转移到静水压力机 (5406R,大岳,日本制)中,施压650 MPa,抑制卵裂,6 min后移至海水 (17℃)中孵化。自2009年5月起,采用室内水槽养殖方法进行繁殖,于2010年11月采集52尾DH个体进行表型和基因型分析。
1.1.2 引物与试剂 根据GeneBank库中的牙鲆微卫星序列设计并筛选出有效微卫星引物94个。引物由上海生工生物技术服务有限公司合成;Taq DNA聚合酶、dNTPs等生化试剂以及DL2000 DNA Maker均购自北京全式金生物技术有限公司。
1.2 方法
1.2.1 性状测量 使用电子天平称量体质量,精确到0.1 g;用X-射线机 (LX-20A型,朗安公司产品)拍摄牙鲆骨骼的X光照片 (图1)。用Motic Images Plus 2.0软件测量照片上的鱼体性状,获得第10腹椎骨长、骨高等性状,精确到0.01 cm。第10腹椎骨长为该块脊椎骨从牙鲆头侧到尾侧的长度,骨高是与其长垂直的高度。
图1 牙鲆X-射线图
Fig.1 The X-ray photography of Japanese flounder Paralichthys olivaceus
1.2.2 牙鲆基因组DNA的提取 利用牙鲆尾鳍组织提取基因组 DNA,将尾鳍剪碎,加入裂解液[50 mmol/L Tris-HCl,100 mmol/L NaCl,20 mmol/L EDTA(pH 8.0),200 mg/L的蛋白酶K, 1%SDS],使其在50℃条件下完全溶解,用相同体积的饱和酚、氯仿混合液抽提1次,再用等体积的异丙醇沉淀,然后用体积分数75%的乙醇清洗沉淀,待TE将沉淀溶解后,放入冰箱 (-20℃)中保存以备用。
1.2.3 PCR反应 10 μL PCR反应体系包含10× buffer 1 μL、dNTPs 0.6 μL、20 pmol/L上下游引物各0.3 μL、模板 DNA 0.4 μL、Taq DNA聚合酶0.04 μL,ddH2O 7.36 μL。PCR反应程序:94℃下预变性5 min;94℃下变性30 s,50~65℃下退火30 s,72℃下延伸30 s,共进行25个循环;最后在72℃延伸10 min。
1.2.4 微卫星分子标记的检测 PCR扩增产物用8% 非变性聚丙烯酰胺凝胶电泳进行检测,使用1× TBE作为电泳缓冲液,在250 V电压下,根据片段大小电泳3 h左右,电泳后将凝胶放入配制好的染色液 (0.5 g硝酸银,500 mL蒸馏水)中银染10 min,经5~10 min的显色液显色,显色液由5 mL HCHO、10 g NaOH、0.02 g NaCO3和500 mL蒸馏水配制而成。凝胶的染色及显色均在摇床上进行,使其颜色均匀,并将凝胶放入透明塑封袋保存。使用凝胶成像系统对凝胶进行拍照。
1.3 数据处理
采用PopGene 32软件对各微卫星基因座的等位基因数 (Na)、有效等位基因数 (Ne)、等位基因频率 (allele frequency)、群体杂合度 (H)进行统计。多态性信息含量 (PIC)通过PIC_Calc 0.6软件计算得出。
使用SPSS 13.0软件中GML程序对双单倍体牙鲆的体质量、第10腹椎骨长和骨高等性状与94个微卫星位点的相关性进行分析。
2.1 牙鲆性状分布
测量性状皆显示出连续变异的特点,所以这些性状都是典型的数量性状。体质量、第10腹椎的骨长和骨高等性状的平均值、偏度、峰度、最小值、最大值以及P值见表1。
表1 体质量、第10腹椎骨长和骨高的正态分布检验
Tab.1 The Gaussian distribution in body weight,and the tenth vertebral length and height
性状traits 平均值±标准误mean±σ 偏度skewness 峰度kurtosis 最小值minimum 最大值maximum P体质量body weight/g 919.227±57.527 0.503 -0.220 279.000 2013.800 <0.05第10腹椎骨长the tenth vertebral length/cm 0.763±0.016 -0.040 -0.178 0.485 0.990 <0.05第10腹椎骨高the tenth vertebral height/cm 0.909±0.019 -0.382 -0.396 0.550 1.150 <0.05
2.2 微卫星基因位点的统计信息
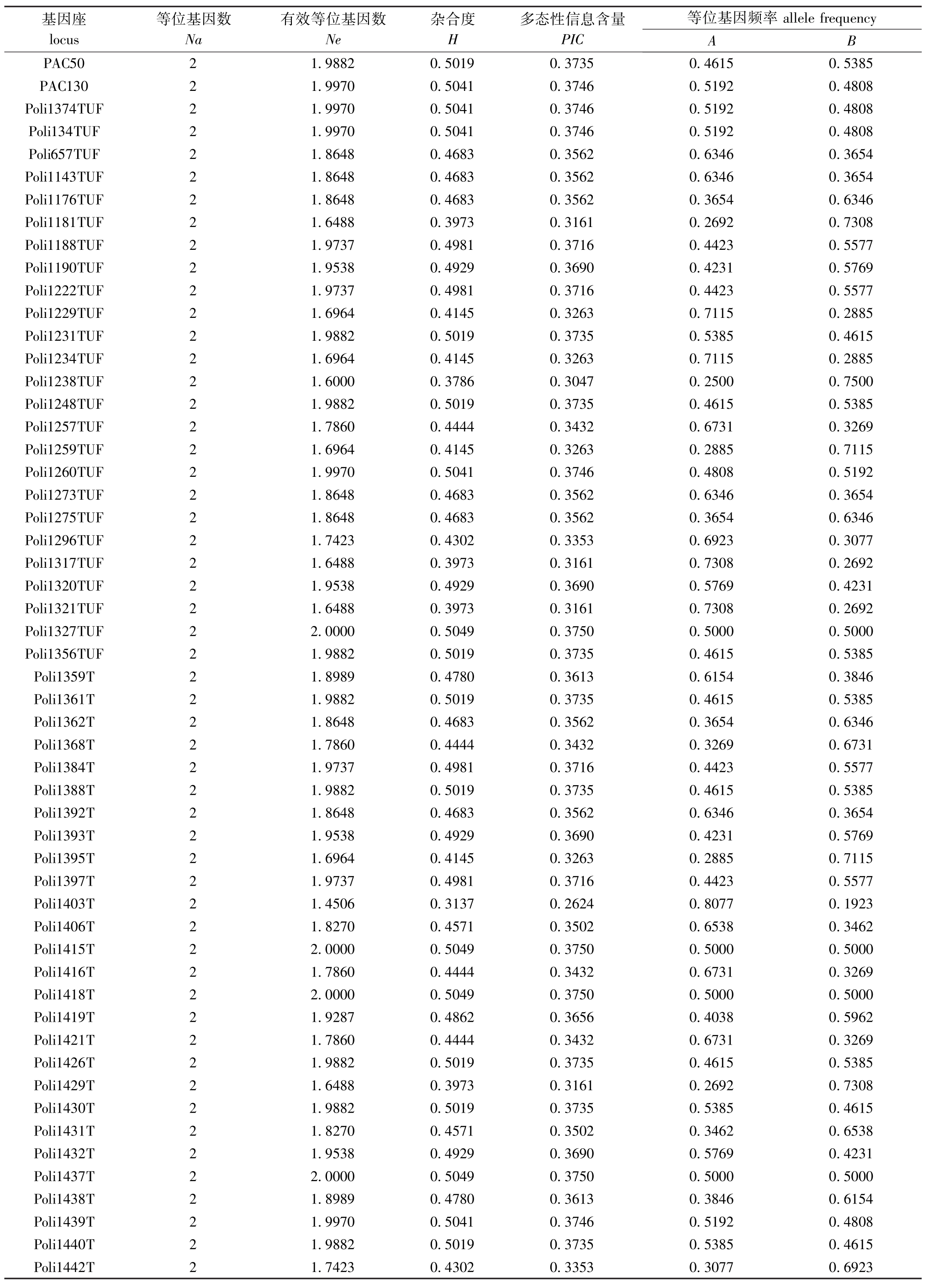
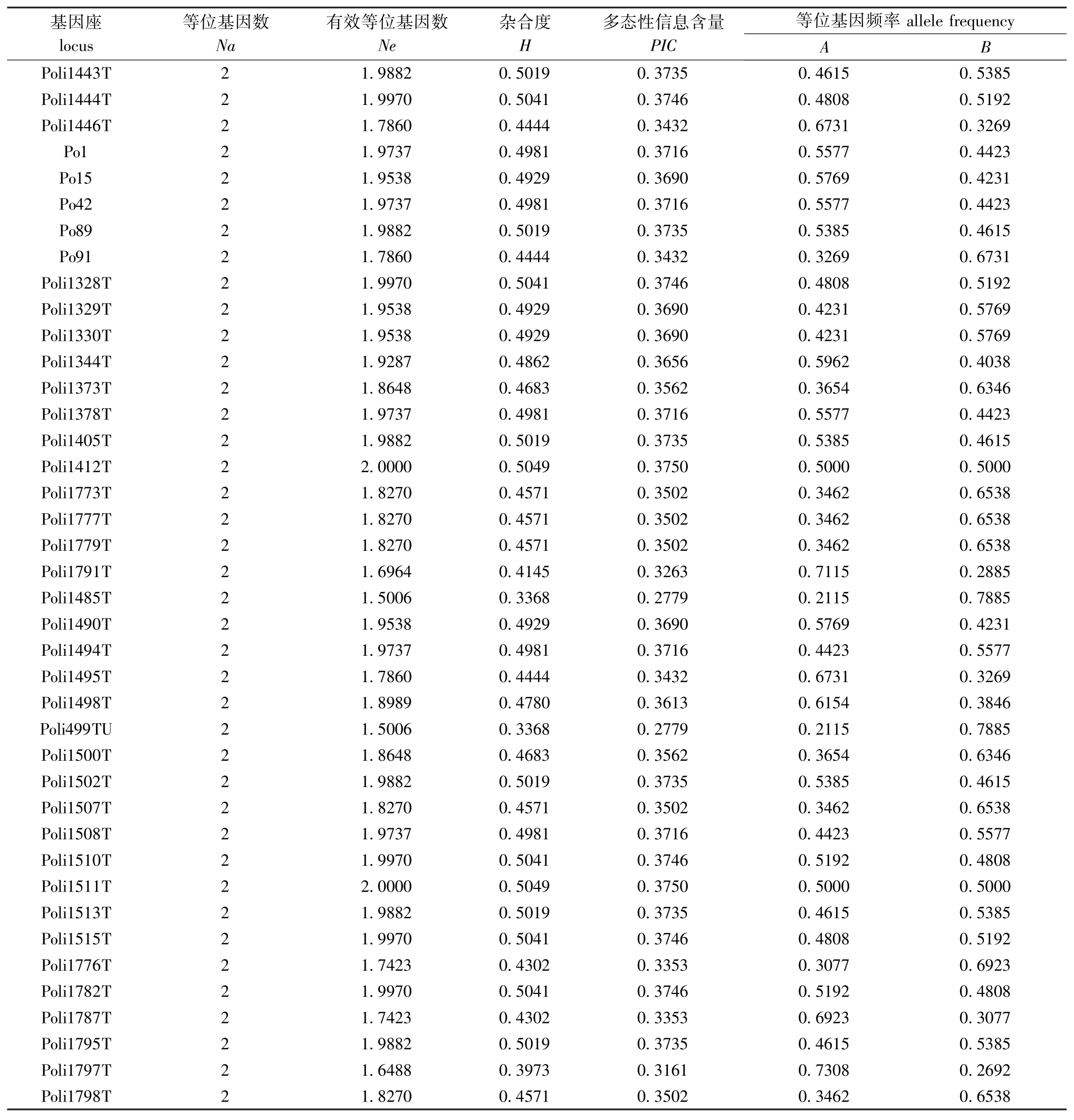
检测94对微卫星标记,共获得188个等位基因。平均Ne为1.878 3,平均H为0.469 2,平均PIC为0.355 8(表2)。
表2 群体在94个微卫星标记基因位点的统计信息
Tab.2 Statistics information of 94 microsatellite genetic loci in the Japanese flounder population
基因座locus等位基因数Na有效等位基因数Ne杂合度H多态性信息含量PIC等位基因频率allele frequency A B PAC50 2 1.9882 0.5019 0.3735 0.4615 0.5385 PAC130 2 1.9970 0.5041 0.3746 0.5192 0.4808 Poli1374TUF 2 1.9970 0.5041 0.3746 0.5192 0.4808 Poli134TUF 2 1.9970 0.5041 0.3746 0.5192 0.4808 Poli657TUF 2 1.8648 0.4683 0.3562 0.6346 0.3654 Poli1143TUF 2 1.8648 0.4683 0.3562 0.6346 0.3654 Poli1176TUF 2 1.8648 0.4683 0.3562 0.3654 0.6346 Poli1181TUF 2 1.6488 0.3973 0.3161 0.2692 0.7308 Poli1188TUF 2 1.9737 0.4981 0.3716 0.4423 0.5577 Poli1190TUF 2 1.9538 0.4929 0.3690 0.4231 0.5769 Poli1222TUF 2 1.9737 0.4981 0.3716 0.4423 0.5577 Poli1229TUF 2 1.6964 0.4145 0.3263 0.7115 0.2885 Poli1231TUF 2 1.9882 0.5019 0.3735 0.5385 0.4615 Poli1234TUF 2 1.6964 0.4145 0.3263 0.7115 0.2885 Poli1238TUF 2 1.6000 0.3786 0.3047 0.2500 0.7500 Poli1248TUF 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1257TUF 2 1.7860 0.4444 0.3432 0.6731 0.3269 Poli1259TUF 2 1.6964 0.4145 0.3263 0.2885 0.7115 Poli1260TUF 2 1.9970 0.5041 0.3746 0.4808 0.5192 Poli1273TUF 2 1.8648 0.4683 0.3562 0.6346 0.3654 Poli1275TUF 2 1.8648 0.4683 0.3562 0.3654 0.6346 Poli1296TUF 2 1.7423 0.4302 0.3353 0.6923 0.3077 Poli1317TUF 2 1.6488 0.3973 0.3161 0.7308 0.2692 Poli1320TUF 2 1.9538 0.4929 0.3690 0.5769 0.4231 Poli1321TUF 2 1.6488 0.3973 0.3161 0.7308 0.2692 Poli1327TUF 2 2.0000 0.5049 0.3750 0.5000 0.5000 Poli1356TUF 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1359T 2 1.8989 0.4780 0.3613 0.6154 0.3846 Poli1361T 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1362T 2 1.8648 0.4683 0.3562 0.3654 0.6346 Poli1368T 2 1.7860 0.4444 0.3432 0.3269 0.6731 Poli1384T 2 1.9737 0.4981 0.3716 0.4423 0.5577 Poli1388T 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1392T 2 1.8648 0.4683 0.3562 0.6346 0.3654 Poli1393T 2 1.9538 0.4929 0.3690 0.4231 0.5769 Poli1395T 2 1.6964 0.4145 0.3263 0.2885 0.7115 Poli1397T 2 1.9737 0.4981 0.3716 0.4423 0.5577 Poli1403T 2 1.4506 0.3137 0.2624 0.8077 0.1923 Poli1406T 2 1.8270 0.4571 0.3502 0.6538 0.3462 Poli1415T 2 2.0000 0.5049 0.3750 0.5000 0.5000 Poli1416T 2 1.7860 0.4444 0.3432 0.6731 0.3269 Poli1418T 2 2.0000 0.5049 0.3750 0.5000 0.5000 Poli1419T 2 1.9287 0.4862 0.3656 0.4038 0.5962 Poli1421T 2 1.7860 0.4444 0.3432 0.6731 0.3269 Poli1426T 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1429T 2 1.6488 0.3973 0.3161 0.2692 0.7308 Poli1430T 2 1.9882 0.5019 0.3735 0.5385 0.4615 Poli1431T 2 1.8270 0.4571 0.3502 0.3462 0.6538 Poli1432T 2 1.9538 0.4929 0.3690 0.5769 0.4231 Poli1437T 2 2.0000 0.5049 0.3750 0.5000 0.5000 Poli1438T 2 1.8989 0.4780 0.3613 0.3846 0.6154 Poli1439T 2 1.9970 0.5041 0.3746 0.5192 0.4808 Poli1440T 2 1.9882 0.5019 0.3735 0.5385 0.4615 Poli1442T 2 1.7423 0.4302 0.3353 0.3077 0.6923
续表2 群体在94个微卫星标记基因位点的统计信息
Cont.Tab.2 Statistics information of 94 microsatellite genetic loci in the Japanese flounder population
基因座locus等位基因数Na有效等位基因数Ne杂合度H多态性信息含量PIC等位基因频率allele frequency A B Poli1443T 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1444T 2 1.9970 0.5041 0.3746 0.4808 0.5192 Poli1446T 2 1.7860 0.4444 0.3432 0.6731 0.3269 Po1 2 1.9737 0.4981 0.3716 0.5577 0.4423 Po15 2 1.9538 0.4929 0.3690 0.5769 0.4231 Po42 2 1.9737 0.4981 0.3716 0.5577 0.4423 Po89 2 1.9882 0.5019 0.3735 0.5385 0.4615 Po91 2 1.7860 0.4444 0.3432 0.3269 0.6731 Poli1328T 2 1.9970 0.5041 0.3746 0.4808 0.5192 Poli1329T 2 1.9538 0.4929 0.3690 0.4231 0.5769 Poli1330T 2 1.9538 0.4929 0.3690 0.4231 0.5769 Poli1344T 2 1.9287 0.4862 0.3656 0.5962 0.4038 Poli1373T 2 1.8648 0.4683 0.3562 0.3654 0.6346 Poli1378T 2 1.9737 0.4981 0.3716 0.5577 0.4423 Poli1405T 2 1.9882 0.5019 0.3735 0.5385 0.4615 Poli1412T 2 2.0000 0.5049 0.3750 0.5000 0.5000 Poli1773T 2 1.8270 0.4571 0.3502 0.3462 0.6538 Poli1777T 2 1.8270 0.4571 0.3502 0.3462 0.6538 Poli1779T 2 1.8270 0.4571 0.3502 0.3462 0.6538 Poli1791T 2 1.6964 0.4145 0.3263 0.7115 0.2885 Poli1485T 2 1.5006 0.3368 0.2779 0.2115 0.7885 Poli1490T 2 1.9538 0.4929 0.3690 0.5769 0.4231 Poli1494T 2 1.9737 0.4981 0.3716 0.4423 0.5577 Poli1495T 2 1.7860 0.4444 0.3432 0.6731 0.3269 Poli1498T 2 1.8989 0.4780 0.3613 0.6154 0.3846 Poli499TU 2 1.5006 0.3368 0.2779 0.2115 0.7885 Poli1500T 2 1.8648 0.4683 0.3562 0.3654 0.6346 Poli1502T 2 1.9882 0.5019 0.3735 0.5385 0.4615 Poli1507T 2 1.8270 0.4571 0.3502 0.3462 0.6538 Poli1508T 2 1.9737 0.4981 0.3716 0.4423 0.5577 Poli1510T 2 1.9970 0.5041 0.3746 0.5192 0.4808 Poli1511T 2 2.0000 0.5049 0.3750 0.5000 0.5000 Poli1513T 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1515T 2 1.9970 0.5041 0.3746 0.4808 0.5192 Poli1776T 2 1.7423 0.4302 0.3353 0.3077 0.6923 Poli1782T 2 1.9970 0.5041 0.3746 0.5192 0.4808 Poli1787T 2 1.7423 0.4302 0.3353 0.6923 0.3077 Poli1795T 2 1.9882 0.5019 0.3735 0.4615 0.5385 Poli1797T 2 1.6488 0.3973 0.3161 0.7308 0.2692 Poli1798T 2 1.8270 0.4571 0.3502 0.3462 0.6538
2.3 第10腹椎骨长、骨高与体质量的相关性分析
利用最小二乘法对第10腹椎的骨长、骨高与体质量进行连锁显著性检验,第10腹椎的骨长、骨高与体质量的相关系数分别为0.836、0.871,相关系数均达极显著水平 (P<0.01)。
2.4 牙鲆微卫星标记与体质量、第10腹椎骨长和骨高的相关性分析
利用最小二乘法,对微卫星标记与牙鲆体质量、第10腹椎骨长和骨高进行相关性检验,在94个微卫星位点中,微卫星标记 Poli1222TUF、Poli1418TUF、Po1、Po91、Poli1498TUF、Poli11795-TUF与第10腹椎骨长显著相关 (P<0.05);Poli1406TUF与第10腹椎骨高显著相关 (P<0.05); Poli1143TUF与第10腹椎骨长和骨高均显著相关(P<0.05);Poli1356TUF和 Poli1388TUF与体质量、第10腹椎骨长和骨高均显著或极显著相关(P<0.05,P<0.01)(表3)。
表3 10个微卫星位点不同基因型体质量、第10腹椎的骨长与骨高的平均值
Tab.3 Means of body weight,and the tenth vertebral length and height in the 10 microsatellite loci

注:*表示差异显著(P<0.05);**表示差异极显著(P<0.01)。
Note:*represents significant difference(P<0.05);**represents very significant difference(P<0.01).
座位locus基因型genotype个体数number体质量/g body weight第10腹椎骨长/cm the tenth vertebral length第10腹椎骨高/cm the tenth vertebral height Poli1143TUF AA 33 966.421±75.348 0.770±0.019*0.923±0.025*BB 19 837.258±86.858 0.752±0.029*0.885±0.030*Poli1222TUF AA 23 890.430±81.948 0.740±0.024*0.912±0.029 BB 29 942.066±81.127 0.782±0.021*0.908±0.026 Poli1418TUF AA 26 948.038±91.402 0.787±0.023*0.904±0.031 BB 26 890.415±71.286 0.740±0.020*0.915±0.024 Po1 AA 29 972.634±76.527 0.780±0.022*0.933±0.226 BB 23 851.887±87.047 0.746±0.023*0.880±0.032 Po91 AA 17 854.171±96.453 0.757±0.032*0.896±0.032 BB 35 950.826±71.826 0.766±0.018*0.916±0.024 Poli1498TUF AA 32 885.125±76.184 0.760±0.021*0.891±0.025 BB 20 973.790±87.693 0.770±0.024*0.939±0.028 Poli11795TUF AA 24 981.104±91.563 0.791±0.022*0.916±0.026 BB 28 866.189±72.539 0.740±0.021*0.904±0.028 Poli1356TUF AA 24 913.512±80.590*0.759±0.024**0.898±0.027*BB 28 924.121±82.856*0.767±0.021**0.920±0.027*Poli1388TUF AA 24 815.800±75.031*0.725±0.021**0.874±0.029*BB 28 1007.879±82.872*0.796±0.021**0.940±0.024*Poli1406TUF AA 34 953.341±66.580 0.766±0.021 0.937±0.021*BB 18 854.789±109.791 0.758±0.024 0.858±0.035*
双单倍体牙鲆因其所有的基因都已纯合固定,可直接应用于目标性状各种基因组合的分离和固定,缩短育种年限[17]。DH群体可通过雌核发育方式获得后代,而不改变其遗传组成,可多年多点试验,有效减少误差,从而有利于QTL分析和数量性状的选择[18]。本研究中选择的试验材料为牙鲆双单倍体群体,每一个体完全同质纯合,同一位点只有两种等位基因,可以提高在分析标记与性状相关性时的准确性。
动物骨骼受遗传因素的影响较大,而受外环境的影响较小[14-15],骨骼参数是一种相对稳定的指标。牙鲆脊柱与肋骨共同起到支撑和承担鱼体质量的作用。因此,鱼体质量的变化,与脊椎骨的大小关系密切。鱼体质量越大,脊柱所要承担的重量越大,脊椎骨高和骨长就越大。鱼类的脊柱由一连串的脊椎骨所组成,一般分为腹椎和尾椎。脊柱长是构成牙鲆体长的重要组成部分,脊椎骨长与体长有密切联系;鱼体高度则由脊椎骨高和肋骨长组成,脊椎骨的长和高对于牙鲆的体长和体高均有重要影响。已有研究表明,牙鲆的体长、体高与体质量显著相关[16,19]。但迄今尚未见到脊椎骨与体质量相关性的研究报道。牙鲆有腹椎骨10个[20],第10腹椎骨为牙鲆腹椎骨的最后一块,根据鱼体形状、X光片观察和骨骼力学的特征,以最小材料承担最大负荷的特点[21],牙鲆头侧第1腹椎骨到第10腹椎骨,其大小呈逐渐增大趋势,由第11腹椎骨,也就是第1个尾椎骨开始,有逐渐减小趋势。因此,第10腹椎骨为整个脊柱中相对较大的一个,可以作为代表。本试验结果表明,第10腹椎骨长、骨高与体质量的相关系数分别为0.836和0.871,均呈极显著相关,表明第10腹椎骨长和骨高可以作为选育体质量值的指标之一。而找到与此性状相关的微卫星标记,即可在牙鲆幼年时期通过分子手段检测出第10腹椎骨长和骨高这两种表型值大的个体,从而预测体质量大的个体,达到标记辅助选育的目的。
本研究结果表明,Poli1222TUF、Poli1418TUF、Po1、Po91、Poli1498TUF、Poli11795TUF等6个标记与第10腹椎骨长显著相关;Poli1406TUF与第10腹椎骨高显著相关;Poli1143TUF与第10腹椎骨长、骨高显著相关;Poli1356TUF、Poli1388TUF与体质量以及第10腹椎的骨长、骨高均显著或极显著相关。Poli1222TUF和 Poli11795TUF均位于11号连锁群上[22],但是位置较远,Po91和 Poli1143TUF均位于9号连锁群上,且距离较近,说明这两个基因可能共同影响脊椎骨长这个性状, Poli1418TUF、 Po1、 Poli1498TUF、 Poli1406TUF、Poli1356TUF、Poli1388TUF等标记分别位于4、17、22、20、23、5连锁群上,呈分散趋势[22]。达到显著相关的标记与性状有的是一个标记同几个性状相关,有的是几个标记同一个性状相关,可能是一因多效,或者多因一效的现象。尽管这些标记还需要进一步的验证,但其统计结果也可为今后分子标记辅助育种提供重要参考。
参考文献:
[1] 石宇光,刘海金,刘永新.雌核发育牙鲆家系的生长比较和形态分析[J].大连海洋大学学报,2010,25(4):324-329.
[2] William P Y,Paul A W,Virginia H C,et al.A detailed linkage map of rainbow trout produced using doubled haploids[J].Genetics,1998,148(2):839-850.
[3] Sato T,Yokomizo S,Matsuda M,et al.Gene-centromere mapping of medaka sex chromosomes using triploid hybrids between Oryzias latipes and O.luzonensis[J].Genetica,2001,111(1/3):71-75.
[4] 盛岩,郑蔚虹,裴克全.马克平微卫星标记在种群生物学研究中的应用[J].植物生态学报,2002,26:119-126.
[5] 孙效文,梁利群.鲤鱼的遗传连锁图谱(初报)[J].中国水产科学,2000,7(1):1-5.
[6] Knapik E W,Goodman A,Ekker M,et al.A microsatellite genetic linkage map for zebrafish Danio rerio[J].Nature Genetics,1998, 18:338-343.
[7] 周莉,刘静霞,桂建芳.应用微卫星标记对雌核发育银鲫的遗传多样性初探[J].动物学研究,2001,22(4):257-264.
[8] 孙效文,梁利群.斑马鱼SSLP标记检测鲤鱼种间的遗传多态性[J].中国水产科学,2001,8(2):5-6.
[9] 池喜峰,贾智英,李池陶,等.荷包红鲤与德国镜鲤正反杂交组遗传结构的微卫星分析[J].大连海洋大学学报,2010,25(5): 450-455.
[10] Brooker A L,Cook D,Bentzen P,et al.Organization of microsatellites differs between mammals and cold-water teleost fishes[J]. Canadian Journal of Fisheries and Aquatic Sciences,1994,51(9): 1959-1966.
[11] Palti Y,Shirak A,Cnaani A,et al.Detection of genes with deleterious alleles in an inbred line of tilapia Oreochromis aureus[J]. Aquaculture,2002,206(3/4):151-164.
[12] Kocabas A M,Kucuktas H,Dunham R A,et al.Molecular characterization and differential expression of the myostatin gene in channel catfish Ictalurus punctatus[J].Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression,2002,1575(1/3):99-107.
[13] 李文升,刘翠,张晓峰,等.镜鲤体重的QTL定位[J].基因组学与应用生物学,2011,30(4):316-324.
[14] 王秋荣,毕建功,林利民,等.青石斑鱼骨骼发育异常的形态特征[J].大连海洋大学学报,2012,27(5):417-421.
[15] 李九军,李勤,贺永春,等.遗传因素对同卵双生子颅颌面部软硬组织形态的影响研究[J].同济大学学报:医学版,2009, 30(3):70-73.
[16] 王凯,刘海金,刘永新,等.牙鲆形态性状对体重的影响效果分析[J].上海海洋大学学报,2008,17(6):655-660.
[17] Foroughi-Wehr B,Wenzel G.Recurrent selection alternating with haploid steps—a rapid breeding procedure for combining agronomic traits in inbreeders[J].Theoretical and Applied Genetics,1990, 80(4):564-568.
[18] Komen H,Thorgaard G H.Androgenesis,gynogenesis and the production of clones in fishes:A review[J].Aquaculture,2007,269 (1):150-173.
[19] 王桂兴,刘永新,孙效文,等.牙鲆微卫星分子标记与生长性状的相关性分析[J].东北农业大学学报,2009,40(7):77-84.
[20] 李思中,王惠民.中国蝶形目鱼类骨骼的研究Ⅱ:脊椎骨、肋骨、上肋骨及肌隔骨刺[J].动物学报,1987,33(3):267-276.
[21] 杨安峰,程红,姚锦仙.脊椎动物比较解剖学[M].北京:北京大学出版社,2008:85.
[22] Castaňo-Sánchez C,Fuji K,Ozaki A,et al.A second generation genetic linkage map of Japanese flounder(Paralichthys olivaceus) [J].BMC Genomics,2010,11(1):554-564.
Correlation of the tenth vertebra with microsatellite DNA marks in Japanese flounder Paralichthys olivaceus
Abstract:Ninety-four microsatellite markers were applied to analyze the correlation between the tenth vertebral length and height and microsatellite DNA marks in 52 doubled haploids of Japanese flounder Paralichthys olivaceus. Results showed that there was significant correlation between Poli1222TUF,Poli1418TUF,Po1,Po91,Poli1498TUF,and Poli11795TUF and the tenth vertebral length and that there was significant correlation between Poli1406TUF and the tenth vertebral height(P<0.05).The significant and very significant correlation were observed between Poli1143TUF,Poli1356TUF and Poli1388TUF and the tenth vertebral length and height(P<0.05,or P<0.01),which can be used as a reference to assisted selection of Japanese flounder.
Key words:Paralichthys olivaceus;gynogenesis;double haploid;microsatellite marker;vertebra
中图分类号:S917.4
文献标志码::A
文章编号:2095-1388(2013)04-0367-06
收稿日期:2012-12-24
基金项目:国家鲆鲽类产业技术体系 (CARS-50-G2);公益性行业农业科研专项 (200903046);国家 “863”计划项目 (2012AA10A-408-5)