
姜志强,张志明,赵翀,李丞,姜柏桥,孙阳
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023)
摘要:通过对大连海域太平洋鳕Gadusmacrocephalus的繁殖力、卵径分布、性腺指数 (GSI)和肝指数(HSI)进行周年调查,确定出太平洋鳕的繁殖期,分别测定了不同发育期太平洋鳕性腺、肝脏和肌肉的生化组成、能量密度变化,并观察了不同发育期太平洋鳕性腺和肝脏的组织学变化。结果表明:大连海域太平洋鳕的卵巢从12月开始进入第Ⅳ期,至翌年2月份排放;精巢处于Ⅳ、Ⅴ期的时间较长,11月份就有达到成熟的个体;从太平洋鳕肝脏组织学的变化来看,脂滴的体积变化很大,说明在繁殖过程中,肝脏经历了一个脂肪先积累又被大量消耗的过程;太平洋鳕在繁殖过程中,各个组织的能量密度变化较小,但粗蛋白、粗脂肪、粗灰分等成分变化较大。研究表明,太平洋鳕性腺发育的脂肪主要可能来源于肝脏,而肌肉蛋白的下降可能充当了性腺发育中蛋白质的主要来源。
关键词:太平洋鳕;性腺;肝脏;肌肉;营养来源
太平洋鳕Gadusmacrocephalus又名大头鳕,分布于太平洋北部沿岸海域,从北太平洋西南部的黄海,经韩国至白令海峡和阿留申群岛,沿太平洋东海岸的阿拉斯加、加拿大至美国的洛杉矶一带沿海,在中国主要产于黄海北部。太平洋鳕为典型的冷水性海水鱼类[1-2]。
在对鱼类繁殖生物学的研究中,除研究其繁殖习性外,还采用性腺指数 (GSI)、肝指数 (HSI)及组织学变化等研究其性腺成熟过程。Kejesbu等[3-4]对大西洋鳕的研究中,将性腺指数与肝指数的关系,卵的大小,性腺的生化组成和组织学变化联系在一起,系统地研究了大西洋鳕雌性个体的繁殖生物学特征。对于与大西洋鳕同属的太平洋鳕的生物学和形态学已有过一些研究[2,5],但关于太平洋鳕繁殖生物学的系统研究目前尚未见报道。本研究中,作者研究了大连海域太平洋鳕的繁殖特点,从GSI和HSI的关系,能量密度、生化组成和组织学的变化等方面,研究了不同发育期太平洋鳕的性腺发育特点和营养来源,旨在为进一步掌握其繁殖规律,科学地开展太平洋鳕的人工繁殖提供基础资料。
1.1 材料
试验用太平洋鳕逐月采自大连沿海,共采集样品105尾,其中成熟雄性52尾,体长为258~695 mm,体质量为484~5 138 g;成熟雌性36尾,体长为285~670 mm,体质量为206~5 025 g;幼鱼17尾。
1.2 方法
1.2.1 测定与计算方法 对太平洋鳕样本的生物学指标进行测量后,解剖,采集性腺、肝脏、肌肉样品。用米尺来测量体长;用电子天平称量体质量、去内脏的体质量、性腺质量和肝脏质量,精确至0.01 g。根据下式计算性腺指数 (GSI)和肝指数 (HSI):
用发育至Ⅳ期的卵巢统计怀卵量,计数1 g卵巢的卵粒数,然后乘以性腺质量即为绝对怀卵量;绝对怀卵量与去内脏的体质量之比即为相对怀卵量。在显微镜下以目微尺测量第Ⅳ期卵巢卵粒的卵径,每尾测量100粒。
太平洋鳕各组织的生化组成通过概略养分分析法测定。将各组织样品在65℃恒温下烘干24 h以上至恒重,得出样品的水分含量;采用凯氏定氮法测出样品中的氮含量,再乘以6.25,得出样品的粗蛋白含量;采用索氏抽提法,将样品用乙醚在50℃恒温水浴中反复抽提4 h以上至不再有脂类物质浸出,得出样品的粗脂肪含量;将样品在600℃下恒温灼烧3 h以上至恒重,得出样品的灰分含量;采用WGR-1型氧弹式热量计直接测定样品的能量密度。
1.2.2 组织切片的制作和观察 将性腺和肝脏样品用波恩氏液固定24~48 h后,用常规石蜡切片,切片厚度为6μm,再用H.E.染色,在Nikon Eslipse 50i显微镜下观察,用Nikon Coolpix 5400数码相机照相。
1.3 数据处理
采用单因素方差法进行显著性检验,当差异达到显著性水平 (P<0.05)时,采用Duncan's法进行组间多重比较。
2.1 繁殖力和卵径分布
大连海域太平洋鳕的绝对怀卵量为39~285万粒,平均81万粒,通常鱼体越大,怀卵量就越多,鱼体的绝对怀卵量与体质量 (去内脏)之间呈正相关关系 (图1),回归方程为y=0.092x-14.618,R2=0.7718;太平洋鳕的相对怀卵量为 (0.0755± 0.0272)万粒/g(去内脏的体质量)。
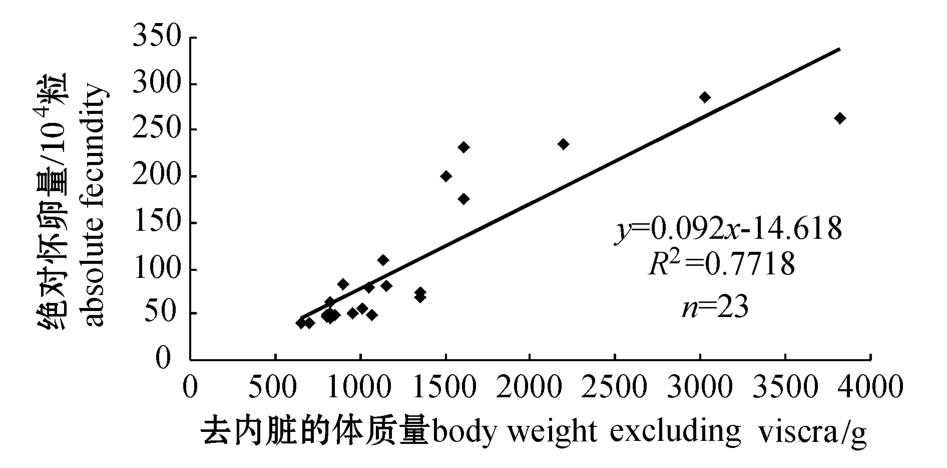
图1 太平洋鳕的绝对怀卵量与去内脏的体质量之间的关系
Fig.1 Relationship between absolute fecundity and
body weight of Pacific cod gutted
12月份太平洋鳕的卵巢已经进入第Ⅳ时相,卵径分布多数集中在450~650μm,呈单峰形(图2)。
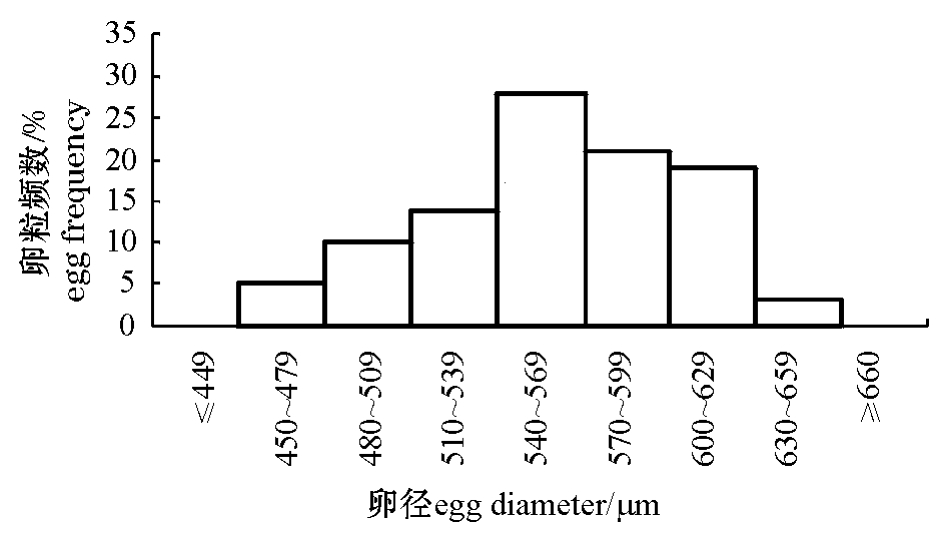
图2 太平洋鳕的卵径分布
Fig.2 Frequency profile in egg diameter of Pacific cod
2.2 性腺指数和肝指数
雌性太平洋鳕的GSI在2月份达到最高值,而HSI在2月份达到最低值 (图3);雄性太平洋鳕的GSI在12月份达到最高值,而HSI在1月份达到最低值 (图4)。
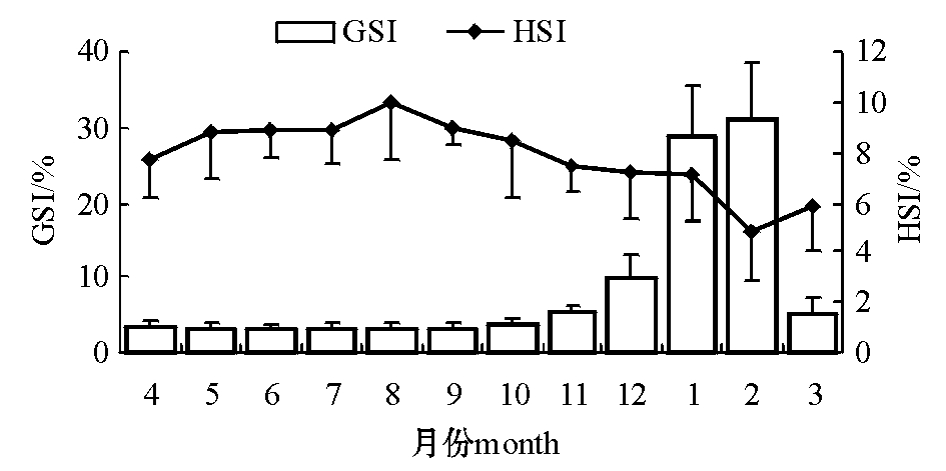
图3 雌性太平洋鳕的性腺指数和肝指数
Fig.3 GSI and HSI of female Pacific cod
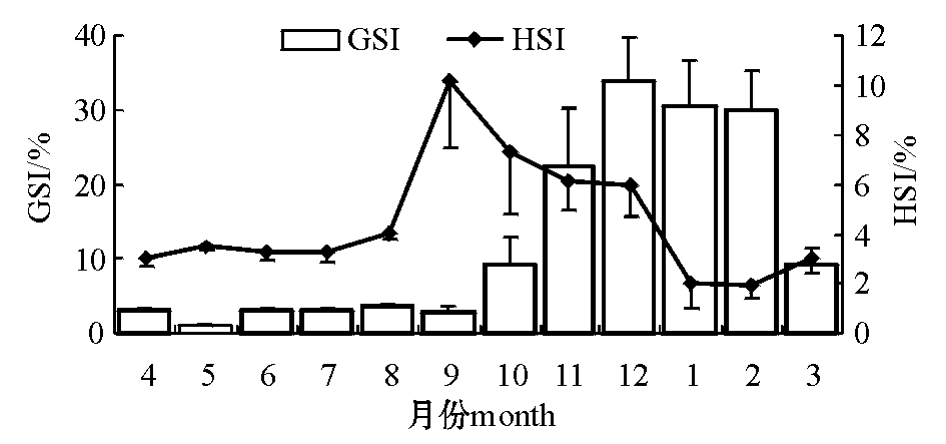
图4 雄性太平洋鳕的性腺指数和肝指数
Fig.4 GSI and HSI ofmale Pacific cod
2.3 性腺、肌肉和肝脏的生化组成和能量密度
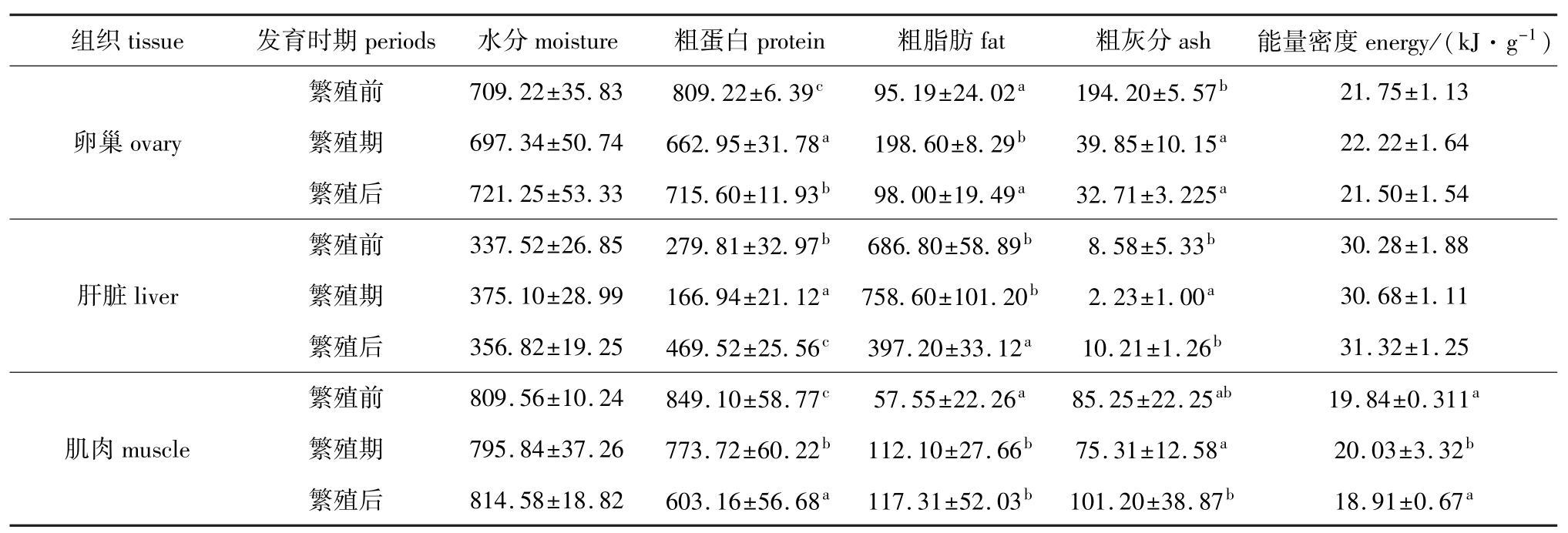
雌性太平洋鳕各个组织的生化组成和能量密度的变化见表1。各组织的粗蛋白、粗脂肪和粗灰分,以及肌肉的能量密度在繁殖前、中、后有较大变化,有的达到差异显著水平 (P<0.05)。其中卵巢中粗蛋白含量繁殖前最高,繁殖期最低,繁殖后又明显较繁殖期增高;肝脏中粗蛋白含量则在繁殖前较高,繁殖期明显降低,繁殖后又显著高于繁殖前和繁殖期;肌肉中粗蛋白含量繁殖前最高,繁殖期明显下降,繁殖后进一步明显下降。卵巢中粗脂肪含量在繁殖前与繁殖后相差不大,但繁殖期明显增高;肝脏中粗脂肪含量繁殖前、繁殖期变化不大,但繁殖后明显降低;肌肉中粗脂肪含量繁殖期和繁殖后相差不大,但都较繁殖前有明显增加。卵巢中粗灰分含量繁殖期和繁殖后相差不大,但都较繁殖前明显下降;肝脏中粗灰分含量繁殖前和繁殖后相差不大,但都明显高于繁殖期;肌肉中粗灰分含量繁殖前与繁殖期、繁殖后相差不大,但繁殖期明显低于繁殖后;除肌肉中能量密度繁殖前、后相近,明显低于繁殖期外,卵巢和肝脏中能量密度在各个期没有明显差别。
雄性太平洋鳕各个组织的生化组成和能量密度的变化见表2。各组织的粗蛋白、粗脂肪、粗灰分均在繁殖前、中、后也有较大变化,有的达到差异显著水平 (P<0.05),但能量密度都变化不大。其中精巢中粗蛋白含量繁殖期和繁殖后相差不大,均明显高于繁殖前;肝脏中粗蛋白含量繁殖前和繁殖期相差不大,但都明显低于繁殖后;肌肉中粗蛋白含量繁殖前和繁殖期相差不大,但均明显高于繁殖后。精巢中粗脂肪含量在繁殖前与繁殖后相差不大,但繁殖期明显增高;肝脏中粗脂肪含量繁殖前、繁殖期变化不大,但繁殖后明显降低;肌肉中粗脂肪含量繁殖前和繁殖期相差不大,但都明显高于繁殖后。精巢中粗灰分含量繁殖前和繁殖期相差不大,但都明显高于繁殖后;肝脏中粗灰分含量繁殖期明显高于繁殖前,繁殖后又明显高于繁殖期;肌肉中粗灰分含量繁殖前和繁殖期相差不大,均明显高于繁殖后;各组织的能量密度在繁殖前、繁殖期及繁殖后无明显差别。
表1 不同发育时期雌性太平洋鳕各组织的生化组成和能量密度(平均值±标准差)
Tab.1 Chem ical composition and energy in different developmental periods in female Pacific cod(mean±S.D.)g/kg
注:同一列同一组织器官中不同字母表示不同时期差异显著(P<0.05),下同。
Note:Data with different letters in same row and same organs are significantly different,et sequentia.
组织tissue 发育时期periods 水分moisture 粗蛋白protein 粗脂肪fat 粗灰分ash 能量密度energy/(kJ·g-1) 21.75±1.13卵巢ovary 繁殖期 697.34±50.74 662.95±31.78a198.60±8.29b39.85±10.15a22.22±1.64繁殖后 721.25±53.33 715.60±11.93b98.00±19.49a32.71±3.225a21.50±1.54繁殖前 337.52±26.85 279.81±32.97b686.80±58.89b8.58±5.33b繁殖前 709.22±35.83 809.22±6.39c95.19±24.02a194.20±5.57b30.28±1.88肝脏liver 繁殖期 375.10±28.99 166.94±21.12a758.60±101.20b2.23±1.00a30.68±1.11繁殖后 356.82±19.25 469.52±25.56c397.20±33.12a10.21±1.26b31.32±1.25繁殖前 809.56±10.24 849.10±58.77c57.55±22.26a85.25±22.25ab19.84±0.311a肌肉muscle 繁殖期 795.84±37.26 773.72±60.22b112.10±27.66b75.31±12.58a20.03±3.32b繁殖后 814.58±18.82 603.16±56.68a117.31±52.03b101.20±38.87b18.91±0.67a
表2 不同发育时期雄性太平洋鳕各组织的生化组成和能量密度(平均值±标准差)
Tab.2 Chem ical composition and energy in different developmental periods in male Pacific cod(mean±S.D.) g/kg
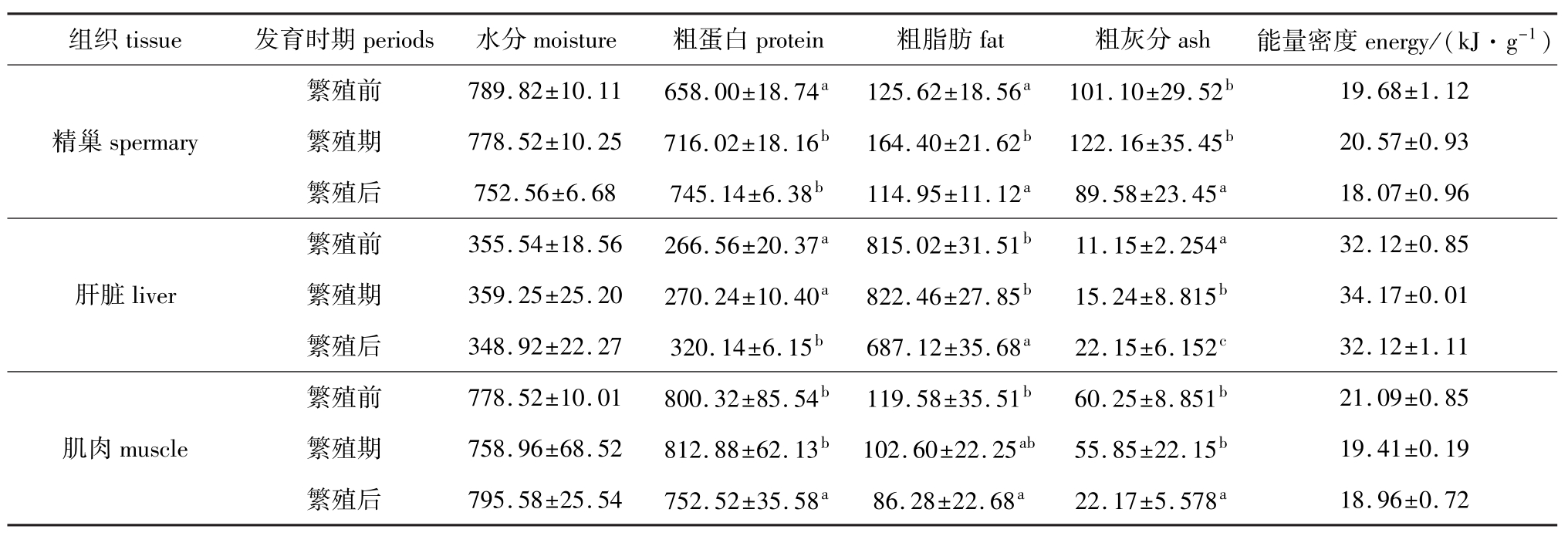
组织tissue 发育时期periods 水分moisture 粗蛋白protein 粗脂肪fat 粗灰分ash 能量密度energy/(kJ·g-1) 19.68±1.12精巢spermary 繁殖期 778.52±10.25 716.02±18.16b164.40±21.62b122.16±35.45b20.57±0.93繁殖后 752.56±6.68 745.14±6.38b114.95±11.12a89.58±23.45a18.07±0.96繁殖前 355.54±18.56 266.56±20.37a815.02±31.51b11.15±2.254a繁殖前 789.82±10.11 658.00±18.74a125.62±18.56a101.10±29.52b21.09±0.85肌肉muscle 繁殖期 758.96±68.52 812.88±62.13b102.60±22.25ab55.85±22.15b19.41±0.19繁殖后 795.58±25.54 752.52±35.58a86.28±22.68a22.17±5.578a32.12±0.85肝脏liver 繁殖期 359.25±25.20 270.24±10.40a822.46±27.85b15.24±8.815b34.17±0.01繁殖后 348.92±22.27 320.14±6.15b687.12±35.68a22.15±6.152c32.12±1.11繁殖前 778.52±10.01 800.32±85.54b119.58±35.51b60.25±8.851b18.96±0.72
2.4 性腺和肝脏的组织学的变化
处于不同发育时期的精巢、卵巢和肝脏的组织学变化见图5。太平洋鳕精巢发育经历了第Ⅱ、Ⅲ、Ⅳ和Ⅴ发育期,Ⅱ期精巢中主要可见精小管,管壁上有大量的精原细胞 (图5-A),Ⅲ期精巢中初级精母细胞充满精小管 (图5-B),Ⅳ期精巢中可见少量的精子游离在精小管腔中 (图5-C),而Ⅴ期精巢中有大量游离的精子存在 (图5-D)。与上述精巢发育期对应的肝脏结构也发生了明显的变化,主要表现在雄性个体肝脏中脂滴体积的增加,Ⅱ期的肝脏脂滴细小,但从Ⅲ期至Ⅴ期,脂滴的体积明显增大 (图5-J、K、L、M)。
太平洋鳕卵巢发育经历了第Ⅱ、Ⅲ、Ⅳ和Ⅴ发育期,Ⅱ期卵巢结缔组织被膜深入,主要是卵黄未形成的初级卵母细胞和少量卵原细胞 (图5-E),Ⅲ期和Ⅳ期卵巢中大生长期的初级卵母细胞和晚期的初级卵母细胞占多数,表现为辐射带形成,卵黄颗粒充满整个核外空间,核膜溶解至消失 (图5-F、G、H),Ⅴ期卵巢中成熟卵排出,只剩下一些残留的卵细胞,滤泡膜脱离,辐射带萎缩 (图5-I)。对应于卵巢各发育期,雌性个体肝脏结构变化规律前期同雄性个体,Ⅱ期肝脏脂滴体积较小,数量少 (图5-N),Ⅲ期和Ⅳ期肝脏中脂滴体积明显增大 (图5-O、P),但从第Ⅳ期到第Ⅴ期的发育中,肝脏中的脂滴被消耗掉,脂滴数量逐渐减少,结缔组织增多,整个结构看上去致密 (图5-Q、R)。
3.1 大连海域太平洋鳕的繁殖特点
与其他地区太平洋鳕的繁殖季节相似,大连地区太平洋鳕的繁殖期为每年的1—2月,即在一年中水温最低的环境下性腺才能达到最终的成熟。据报道,太平洋鳕的产卵场通常在近岸,少数鱼群分散在海州湾外海产卵[2]。但作者认为,由于太平洋鳕产沉性卵,在外海产卵会影响成活率。本研究中繁殖期内的太平洋鳕全部为近岸群体。
大连海域的太平洋鳕卵巢在一年中大约从12月开始进入Ⅳ期,翌年1—2月份达到排放,而精巢11月份就成熟,逐渐有精子排放。图3表明,雌性个体的GSI在繁殖期出现一个明显的高峰, 1—2月份为繁殖期,该期持续时间较长,在此期间都可以获得成熟卵细胞,这与大西洋鳕的繁殖习性一致。
从卵径分布来看,太平洋鳕与大西洋鳕同样属一次产卵类型[6]。另外,从GSI和卵母细胞的发育特征也可以确定太平洋鳕为一次产卵类型,在繁殖期的太平洋鳕卵巢中,成熟卵母细胞占绝对优势,极少见到发育早期的卵母细胞,说明卵母细胞发育是同步的。
大连海域太平洋鳕的绝对怀卵量为39~285万粒,与其他海域相比繁殖力较低。北美地区和白令海中的太平洋鳕,绝对怀卵量平均在200万粒左右,而远东地区的太平洋鳕则高达140~640万粒,平均在400万粒左右[6]。这可能与大连海域太平洋鳕个体较小有关。如果从相对繁殖力的角度来看,各个海域的太平洋鳕相差无几。
3.2 太平洋鳕性腺发育的营养来源
太平洋鳕在繁殖过程中,各组织的能量密度变化较小,这是其维持生命特征的基本条件。但从性腺、肝脏和肌肉的生化组成来看,粗蛋白、粗脂肪、粗灰分等成分变化较大 (表1、表2),这种变化与性腺发育有关。性腺和肝脏的组织学研究表明 (图5),雌性太平洋鳕的肝脏结构变化较大,尤其是肝脏中脂滴的体积变化明显,随着太平洋鳕性腺发育的进行,肝脏营养结构经历了一个脂肪先积累又被大量消耗的过程。这个结果与对南方大口鲇[7]和四川境内的鲇[8]以及大西洋鳕[4]的研究结果相同。从图3可以看出,GSI和HSI两者的变化趋势恰好相反,这说明在太平洋鳕卵巢发育处于最佳状态时,即在卵黄积累的高峰期,肝脏脂肪的消耗量最大,两者间的关系在生殖期间表现得尤为突出,这可能是雌性鱼类在繁殖期间摄食明显减少,卵巢发育所需要的能量主要由肝脏提供。而雄鱼则不存在卵黄蛋白原合成的问题,因此,HSI未呈现出明显的季节变化。总之,太平洋鳕GSI和HSI之间存在密切关系,在研究鱼类生长和生殖生理时,应同时考虑这两个指标。
朱洗[9]指出,鱼类体外受精需要大量的生殖细胞,其大生长的过程必须依赖大量的能量,这种能量主要来源于脂肪代谢。他还将鳕属Gadus的鱼类归结为瘦型鱼,这类瘦型鱼的特点可以描述为肌肉永是瘦削无油,脂肪集中于肝脏;在前成熟阶段肝脏生长快;等到生殖腺急剧大生长时期,肝脏的脂肪性高能量物质一部分移用于滋养生殖腺,另一部分用作雌鱼生殖时其他的急需。所以,HSI的高峰一定出现在GSI之前。本研究结果与此相同。
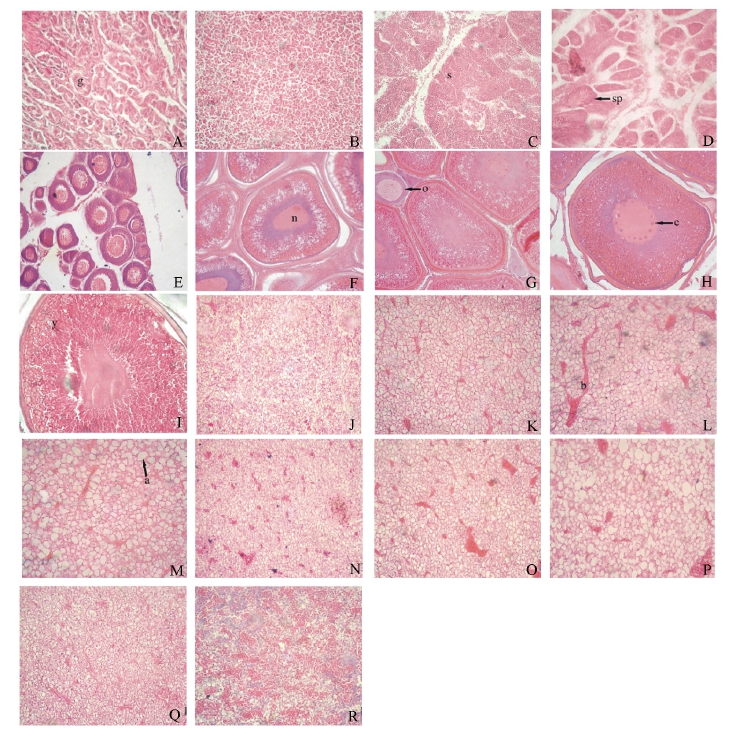
图5 不同发育期太平洋鳕性腺和肝脏组织学的变化
Fig.5 H istological observation of gonad and liver at different stages in Pacific cod
注:AⅡ期精巢,精小管及精原细胞,g示精小管 (×100);BⅢ期精巢,精小管中的初级精母细胞 (×200);CⅣ期精巢,精小管壁上的精子细胞,s示精子细胞 (×200);DⅤ期精巢,成熟精子,sp示精子 (×200);EⅡ期卵巢中小生长期的初级卵母细胞 (×100);FⅢ期卵巢,n示细胞核 (×100);GⅢ期卵巢,o示卵母细胞 (×100);HⅣ期卵巢,c示核仁 (×200);IⅤ期卵巢中的卵母细胞,y示卵黄 (×200);JⅡ期精巢时对应的肝脏结构 (×200);KⅢ期精巢时对应的肝脏结构 (×200);LⅣ期精巢时对应的肝脏结构,b示肝血窦 (×200);MⅤ期精巢时对应的肝脏结构,a示脂滴 (×200);NⅡ期卵巢时对应的肝脏结构 (×200);OⅢ期卵巢时对应的肝脏结构 (×200);PⅣ期卵巢时对应的肝脏结构 (×200);QⅣ期末卵巢时对应的肝脏结构 (×200);RⅤ期卵巢时对应的肝脏结构 (×200)。Note:A StageⅡin testis,seminiferous tubule and spermatogonia,g showing spermatogonia(×100);B StageⅢin testis,primary spermatocytes in seminiferous tubule(×200);C StageⅣin testis,spermatids on the wall of seminiferous tubule,s showing spermatids(×200);D stageⅤin testis,sperm,sp showing sperm(×200);E StageⅡin ovary,primary oocyte in early,growing stage(×100);FStageⅢin ovary,n showing nuclear (×100);G StageⅢin ovary,o showing oocyte(×100);H StageⅣin ovary,c showing nucleolus(×200);IStageⅤin ovary,oocyte,y showing yolk(×200);JLiver structure during stageⅡ in testis(×200);K Liver structure during stageⅢ in testis(×200);L Liver strcture during stageⅣin testis,b showing sinusoid(×200);M Liver structure during stageⅤin testis,a showing fat vacuole(×200);N Liver structure during stageⅡin ovary(×200);O Responding liver structure during stageⅢ in ovary(×200);P Responding liver structure during stageⅣ in ovary (×200);Q Responding liver structure during late stageⅣ in ovary(×200);R Responding liver structure during stageⅣ in ovary(×200).
北冰洋中的极鳕在繁殖期性腺指数、肝指数、肝脏脂肪和肌肉蛋白质都发生了较大变化[10],在挪威的大西洋鳕也出现了同样的趋势[11]。Rideout等[12]的研究表明,卵巢闭锁的大西洋鳕性腺发育的营养来源有所分化,脂肪来源于肝脏,蛋白质来源于肌肉,对于不再继续发育的卵巢,其肝脏也同时停止变化。Love[13]在对大西洋鳕的研究中,还发现这种营养的变化受温度的影响,或者说温度变化的刺激决定了其发育。这些研究结果虽然不尽相同,但大多都把性腺发育的营养来源归结于肝脏和肌肉上。本研究中,太平洋鳕也存在类似的现象,且性腺、肝脏、肌肉的脂肪和蛋白质的变化都达到了显著性水平。太平洋鳕肝脏中脂肪在繁殖后显著下降,可能已转移用于性腺发育和繁殖,繁殖后肌肉中蛋白质显著下降可能充当了性腺发育中蛋白质的主要来源。
参考文献:
[1] 张春霖,成庆泰,郑葆珊,等.黄渤海鱼类调查报告[M].北京:科学技术出版社,1955.
[2] 高天翔,张肖荣,王丹,等.几种鳕鱼的生物学初步研究[J].海洋湖沼通报,2003(1):35-42.
[3] Kjesbu O S,Solemda P,Bratland P,et al.Variation in annual egg production in individual captive Atlantic cod(Gadusmorhua) [J].Can JFish Aquat Sci,1996,53:610-620.
[4] Kjesbu O S,Klungsoyr J.Fecundity,atresia,and egg size of captive Atlantic cod(Gadusmorhua)in relation to proximate body composition[J].Can JFish Aquat Sci,1991,48:2333-2343.
[5] 高天翔,武云飞,张秀梅,等.四种鳕鱼的形态学研究[J].青岛海洋大学学报,2002,32(6):884-896.
[6] Cohen DM,lnada T,lwamoto T,et al.Food and agriculture organization of the United Nations:Rome,Gadiform Fishes of the World (Order Gadiformes)[C]//An Annotated and Illustrated Catalogue of Cods,Hakes,Grenadiers and other Gadiform Fishes Known to Date,1990:18-88.
[7] 张耀光,谢小军.南方鲇的繁殖生物学:性腺发育及周年变化[J].水生生物学报,1996,20(1):8-16.
[8] 魏刚,黄林.鲇繁殖生物学研究[J].水产学报,1997,21(3): 225-232.
[9] 朱洗.鱼类的生殖及子代的发育、生长与变态[J].北京:科学出版社,2000:68-71.
[10] Brawn V M.Reproductive behaviour of the cod(GaduscallariasL.)[J].Behaviour,1962,18:177-198.
[11] Ellassen JE,Vahl O.Seasonal variations in biochemical composition and energy content of liver,gonad and muscle ofmature and immature cod,Gadusmorhua(L.)from Balsfjorden,northern Norway[J].JFish Biol,1982,20:703-709.
[12] Rideout R M,Burton M PM,Rose G.Observations on mass atresia and skipped spawning in northern Atlantic cod from Smith Sound,Newfoundland[J].Journal of Fish Biology,2000,57:1429 -1440.
[13] Love R M.Themeasurement of condition in North Sea cod[J].J Cns Int Explor Mer,1962,27:34-42.
The gonad development and nutrition source of Pacific cod Gadusmacrocephalus
JIANG Zhi-qiang,ZHANG Zhi-ming,ZHAO Chong,LICheng,JIANG Bai-qiao,SUN Yang
(Key Laboratory of Mariculture&Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China)
Abstract:Fecundity,egg diameter frequency distribution,gonosomatic index(GSI)and hepatosomatic index (HIS)of Pacific codGadusmacrocephalusin Dalian sea area were investigated yearly in this study.Changes in chemical compositions,energy density and histology of gonad,liver and muscle of Pacific cod were observed and measured in different stages.The ovary of Pacific cod was inⅣstage from December and ovulated in February in the next year.The spermary lasted a long time inⅣand V stages,and with sperm present from November.According to the histological changes in the liver,oil dropswere found to be changed a lot,indicating that liver underwent a process of lipid accumulated and consumed in breeding process.The energy in each tissuewas less influenced in the breeding process,which were the conditions of basic living need.However,protein,fat and ash were changed a lot,which associated with breeding.The results showed that themain lipid source for the gonad development of Pacific codmight come from liver.Themain protein source for the gonad developmentof Pacific cod is suggested to come from muscle.
Keywords:Gadusmacrocephalus;gonad;liver;muscle;nutrition source
中图分类号:S917.4
文献标志码:A
文章编号:2095-1388(2012)04-0315-06
收稿日期:2012-06-12
基金项目:国家 “863”计划项目 (2012AA10A413)
作者简介:姜志强 (1960-),男,教授。E-mail:zhqjiang@dlou.edu.cn