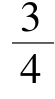 周(图1-b)。
周(图1-b)。施炜纲,徐东坡,刘凯,段金荣,张敏莹
(中国水产科学研究院淡水渔业研究中心,江苏无锡214081)
摘要:采用Pixera Penguin 600CL数码照像机对大银鱼ProtosaIartx hyalocranius胚胎发育的各个时期及仔鱼的开口饵料、仔鱼养殖过程中的摄食行为进行了观察。结果表明:大银鱼受精卵在4~9℃下孵化全程约需852 h,胚胎发育经历了9个生理阶段共30个发育期;出膜后仔鱼平均体长为4.76 mm,全长为4.86 mm;出膜后约20 d卵黄完全吸收;平游期鱼苗平均体长为5.85 mm,全长为6.09 mm。在显微镜下对不投饵养殖条件下的仔鱼进行观察,发现仔鱼早期(10~20 d)以浮游植物为开口饵料,20~40 d时可主动摄取轮虫,仔鱼对轮虫壳及休眠卵无法消化。
关键词:大银鱼;胚胎发育;仔鱼;开口饵料
大银鱼ProtosaIartx hyalocranius主要分布于渤海、黄海、东海沿岸、长江、淮河中下游河道、湖泊和水库中,在中国已形成一定规模产量,且具有较高的经济价值。由于大银鱼适应性强,一直以来是国内天然水域增殖、移植的重要种类。至今绝大多数省市均移植了大银鱼,并获得了成功。
许多学者对大银鱼胚胎发育进行过研究[1-2],但鉴于显微摄影设备及技术条件所限,对大银鱼胚胎发育各期图片的截取及描述尚欠详尽。亦有学者[3]对影响大银鱼孵化率的生态因子开展了研究,有关大银鱼的池塘养殖也有报道[4],但均停留在试验阶段,大银鱼生产性养殖至今尚未突破。对孵出的仔鱼在人工养殖条件下的食性及行为观察亦不够深入且尚未形成共识[5]。本试验中,作者对大银鱼各期胚胎发育及出膜仔鱼习性进行了显微观察与研究,期望能为大银鱼人工孵化及仔鱼培育提供有益的技术参考。
1.1 材料
大银鱼成熟亲体采自太湖三山岛水域,取雌、雄亲体各一尾,体长分别为16、19 cm,以干法人工授精获取受精卵共1.86万粒[6],加注太湖湖水约3.5 L,带回实验室观察。
1.2 方法
孵化养殖用玻璃水族箱(0.8 cm×0.8 cm),采用提前45 d曝气的自来水,保持水位约40 cm。受精卵至原肠后期带水移入箱内,在室温4~9℃条件下孵化。每天取30粒卵在显微镜下观察。
采用Pixera Penguin 600CL荧光显微数码摄像机跟踪拍摄胚胎发育过程,设置每1 h截拍发育图一张,供胚胎发育各期分析。
出膜后仔鱼仍在原孵化水族箱内进行培养,期间不投饵,微充气,观察仔鱼的摄食与活动,每天取30尾仔鱼进行肠道镜检,观察大银鱼开口时机及其天然开口饵料的组成。
2.1 受精与孵化过程
经测定,50粒受精卵平均卵径为1.1 mm,孵化耗时852 h,约35.5 d,至试验结束共截获胚胎发育及开口食性图997张。经筛选分析,大银鱼胚胎发育有9个发育阶段,含30期(表1,图1-A~Z、a~d)。出膜后仔鱼体长为4.76 mm,全长为4.86 mm;平游期鱼苗体长为5.85 mm,全长为6.09 mm。
2.2 胚胎发育
成熟的大银鱼卵近圆球形,沉性,受精卵透明。受精孔(卵膜孔)呈锥形漏斗状,与宋慧春等[7]在电镜下的观察描述一致(图1-A,图2)。卵表面具有细密的卵膜丝,在赤道偏动物极一侧呈圆环状排列。卵膜丝具有较强的吸附力,质量好的受精卵大多可吸附在其他物体上。一旦卵膜丝受损,卵就完全失去黏性,随水流漂移。
2.2.1 胚盘形成 受精后3.5 h,在质膜与卵膜之间出现卵周隙,卵黄粒开始沉积于植物极,原生质逐渐向动物极集中,胚盘形成,单细胞隆起(图1 -A,图2)。
2.2.2 分裂期 在胚盘顶部出现经裂沟,受精卵进入2细胞期。此后开始进行N次卵裂,每次卵裂均垂直于前一次卵裂沟,细胞数以2的N次方增殖,受精卵依次进入32细胞期、64细胞期、128细胞期、256细胞期、多细胞各期。受精后45 h进入桑葚期,此阶段细胞颗粒仍很清晰,此后受精卵分裂速度渐渐变慢(图1-B~H)。
2.2.3 囊胚期 细胞继续分裂,细胞数不断增多,单个细胞体积不断趋小。内层细胞出现腔隙,受精卵进入囊胚早期。随着囊胚继续发育,细胞变得细小,外层细胞颗粒界面变得模糊,囊胚高度逐步降低,受精卵依次进入囊胚中期、囊胚晚期,此时外胚层形成(图1-I~K)。
2.2.4 原肠期 随着囊胚层逐渐下包,胚层变薄变长,受精卵进入原肠早期。胚层各部高度无明显差异,进入原肠中期后,原肠胚略显垂直拉长状,胚层两端略有增厚。随着囊胚下包,只剩下少量卵黄没有被包入,原肠胚又重新趋于圆形(图1-L~N)。
2.2.5 神经胚期 胚体轮廓初现,背侧神经物质增厚,神经沟开始出现,胚体渐变为水平向拉伸,已初具胚体雏形,但头、尾部尚难分辨。受精后约224 h,胚孔封闭,此时头尾分化已十分明显,胚沟延伸一线已无法见到受精孔。受精258 h后进入胚体形成期,胚体形成并又渐变呈圆形,头部器官分化征兆出现,神经管前端分化为前脑、中脑和菱脑三部分。(图1-O~Q)。
2.2.6 器官形成与组织分化阶段 器官形成与组织分化在胚胎发育至289~501.25 h阶段内完成,在头部器官分化初期,体节逐渐形成,从眼囊、嗅板的出现,至尾芽期胚体延伸恰好一周,卵黄囊被挤压成扇形状。随着耳囊及晶体各期的出现,在卵黄囊上方及耳囊下方出现心脏原基,心脏扩大,心包腔清晰,此时体节增至33~40对(图1-R~W)。
2.2.7 胚体蠕动与心脏搏动阶段 此时胚体延伸已近一周半,胚胎开始出现蠕动,初期蠕动幅度不大,频率较低,约6次/h。心脏尚无搏动迹象,胚胎发育到602.5 h,心脏开始缓慢而轻微地搏动,初期心脏搏动频率为6~7次/mim,至638 h耳囊中耳石形成,肌节已达56~64对。此时心脏搏动开始加快,胚体蠕动频繁,间隔3~4 min,幅度也变大,有时能见到整个胚体剧烈蠕动,接连2~3次,胚体与卵膜已无粘连,此时心脏搏动达30次/min(图1-X~Z、a)。
2.2.8 孵化腺出现阶段 卵黄囊缩小为梨形,头部下端稍高于卵黄囊,可见口孔,尾的较大部分已经从卵黄囊分开。胚体可全身翻转,心跳有力,心脏搏动达50~62次/mim,此时胚体环卵约1周(图1-b)。
2.2.9 仔鱼出膜阶段 胚体心跳加速(70~86次/mim),翻转非常剧烈,即进入出膜前期(图1 -c1)。在孵化腺分泌物溶解卵膜及胚体剧烈翻转作用下,卵膜破裂,含脂滴的卵内溶物渗出卵膜(图1-c2)。至835 h仔鱼出膜开始,出膜方式以头部破膜最为多见(图1-d1、d2),也有以尾部破膜或头尾同时破膜的,或以背、胸、腹等部位破膜的,但较少见。刚出膜的仔鱼全长不足5 mm,鳍尚未出现,仅在背部、腹部和尾部有皮肤褶皱。腹部具黑色花纹色素,仔鱼具趋光性。至852 h,进入出膜高峰。至874 h,仔鱼出膜全部完成。出膜后仔鱼平均体长为4.76 mm,平均全长为4.86 mm。初孵仔鱼卵黄囊呈卵圆形。
2.3 大银鱼仔鱼早期的习性
水温为7~10℃时,仔鱼平游需10 d,至平游期鱼苗平均体长为5.85 mm,平均全长为6.09 mm。但至平游后卵黄囊仍未完全吸收,此阶段卵黄囊逐渐转成前粗后细长条形,附于肠管下方,后期卵黄囊长约占体长的60%,卵黄囊腹面每侧可见一列黑色素细胞。仔鱼游动活泼,趋光,此时虽已能平游,但游泳能力仍很弱,只能作短距离平直游动,而后还会下沉,这一阶段可持续20 d左右。
出膜后10~20 d,养殖水体中浮游植物生物量为5.8~6.2 mg/L。在镜下观察仔鱼肠道,发现浮游植物出现率高低依次为:菱形藻、栅列藻、针杆藻、衣藻、裸藻及微囊藻6个属种(图3)。这一时期是仔鱼口器逐步发育完善的阶段,仔鱼在此阶
段的后期摄食行为很特别。仔鱼缓慢游动至适口饵料的下方,头部向上,鱼体收缩成“S”状,逐步靠近目标后,通过身体迅速弹伸向上来捕获目标。
表1 大银鱼胚胎发育的过程
Tab.1 The embryonic development of large icefish

发育期Developmental stage各发育期简要特征Brief characteristics of different stages受精后时间Time after fertilization图号Fig.No. 1细胞期1 cell stage原生质集中于动物极一端,动物极中心处,受精孔可见(俯视),胚盘顶部凸起,透过隆起部在动物极基部上方受精孔清晰可见(侧视) 3.50 h图1-A,图2 2细胞期2 cell stage胚盘顶部中轴线出现卵裂沟,纵向经裂出2个等大的耳状细胞(俯视),2个细胞隔裂线处呈股沟状,受精孔仍可见(侧视)7.00 h图1-B1、B2 4细胞期4 cell stage与第一次卵裂沟相垂直,纬裂为4个等大的扇状细胞(俯视),可见2~3个细胞相错隆起,并高于2细胞期的隆起度(侧视)7.75 h图1-C1、C2 8细胞期8 cell stage每个细胞发生经裂,形成2排4列共8个排列整齐的细胞,胚盘边缘的分裂细胞略小于中央的细胞(俯视)9.16 h图1-D 16细胞期16 cell stage每个细胞发生纬裂,形成4排4列共16个分裂细胞,排列整齐,细胞形状呈正矩形或三角形(俯视)11.33 h图1-E 32细胞期32 cell stage经裂形成32个分裂球,排列欠整齐,细胞形状不规则。细胞堆挤在一起(俯视) 13.58 h图1-F 64细胞期64 cell stage胚盘完成第6次分裂,纬裂成64个分裂球,细胞显著变小,相互重叠14.74 h图1-G桑葚期Multi-cellular经过128、256细胞期及更多期的细胞分裂,细胞呈颗粒状,界面不清(俯视),细胞颗粒团形似桑葚球,高高隆起,与卵黄连接的两侧边缘略有凹陷(侧视) 45.00 h图1-H1、H2囊胚早期Early blastula囊胚高而集中,呈高帽状,与卵黄连接的两侧边缘平滑60.16 h图1-I囊胚中期Mid blastula囊胚层变低、宽,呈月牙形,并沿卵黄的表面向植物极扩展82.83 h图1-J囊胚后期Late blastula囊胚层变得更低、更宽,开始下包卵黄107.00 h图1-K原肠早期Early gastrula胚环形成,胚层下包卵黄近1/2,胚胎纵向拉长呈长圆形132.00 h图1-L原肠中期Mid gastrula胚层下包卵黄约3/5,胚盘仍呈长圆形160.58 h图1-M原肠后期Late gastrula胚层下包卵黄近趋完整,胚盘水平向拉长近似圆形185.58 h图1-N神经胚期Neural formation胚体水平向拉长呈长圆形,胚胎背部表面中线的细胞增厚形成神经板224.00 h图1-P胚体形成期Segmentation胚体的轮廓清晰,胚体水平向收缩,再次呈圆形。258.00 h图1-Q肌节出现期Appearance of myomere出现1~3对肌节及眼囊原基,头部发育分化开始289.00 h图1-R眼囊期Optic capsule stage眼囊清晰,呈长椭圆形,肌节4~9对330.00 h图1-S嗅板期Olfactory plate appearance眼囊下方出现嗅板,肌节10~16对361.00 h图1-T尾芽期Tail-bud stage胚体伸长,头尾相连交错,尾芽形成,针状晶体可见,肌节17~24对201.00 h图1-O胚孔闭合期Closure of blastopore胚层包裹卵黄,胚孔封闭,卵黄腔出现,眼囊隐约可见,胚体呈长圆形391.50 h图1-U耳囊出现期Otocyst stage眼囊凹陷呈杯状,后脑两侧出现小泡状耳囊,肌节25~32对452.75 h图1-V晶体出现期Lense stage眼杯中晶体已清晰可见,肌节33~40对501.25 h图1-W肌肉效应期Muscular contraction胚体环绕卵黄囊一周半呈蛇形盘曲,经常出现扭动,肌节41~48对557.50 h图1-X心脏出现期Appearance of heart卵黄呈扇形,耳囊后下方出现心脏原基,肌节49~56对602.50 h图1-Y耳石出现期Appearance of statolith耳囊出现小石粒,胚体中脑膨大,眼球黑色素沉着,肌节56~64对638.00 h图1-Z心跳期Heart-beating stage心脏跳动有力,心搏率为30次/min,肌节65~66对,肌节由胸背部至尾部逐步趋小672.00 h图1-a卵化腺期hatching gland眼部黑色素沉着明显,头部及体表出现孵化腺,尾部已与卵黄囊分开720.00 h图1-b出膜期Hatching胚体翻转剧烈,卵内溶物渗出,以头部破膜居多852.50 h图1-c1、c2、d1、d2
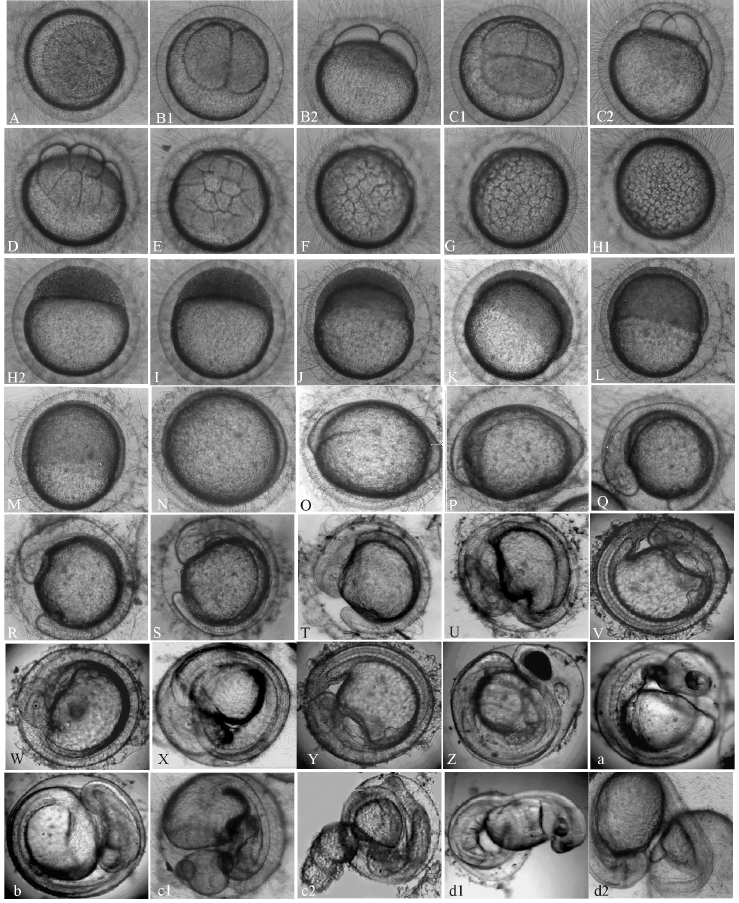
图1 大银鱼的胚胎发育
Fig.1 The embryonic development of large icefish
注:A 1细胞(俯视);B1 2细胞(俯视);B2 2细胞(侧视);C1 4细胞(俯视);C2 4细胞(侧视);D 8细胞(俯视);E 16细胞(俯视);F 32细胞(俯视);G 64细胞(俯视);H1桑葚期(俯视);H2桑葚期(侧视);I囊胚早期;J囊胚中期;K囊胚后期;L原肠早期;M原肠中期;N原肠后期;O神经胚期;P胚孔闭合期;Q胚体形成期;R肌节出现期;S眼囊期;T嗅板期;U尾芽期;V耳囊出现期;W晶体出现期;X肌肉效应期;Y心脏出现期;Z耳石出现期;a心跳期;b孵化腺期;c1出膜前期;c2出膜前期(脂溶物外渗);d1出膜(头部破膜);d2出膜(尾部尚在卵膜内)。
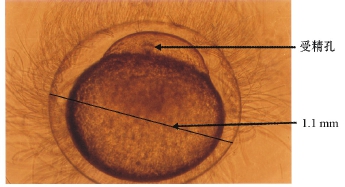
图2 受精卵单细胞胚盘
Fig.2 Disc stage at single-cell stage
图3 肠道内的浮游植物
Fig.3 Phytoplankton in the intestine of the larvae
仔鱼出膜21~40 d内,已完全能平游,营水平游动捕食,全长平均为7.22 mm。口已张开,活泼游动,胸鳍基对体轴几乎呈垂直状态,其下面可见肩带原基。鳔已充气,肝脏形成,约占据5个肌节范围,镜检可见肠管蠕动,能够消化食物。此阶段对肠道进行镜检,发现食物以轮虫为主,按出现率高低依次为:角突臂尾轮虫、蒲达臂尾轮虫、萼花臂尾轮虫、多肢轮虫、三肢轮虫、裂足轮虫、疣毛轮虫。鱼苗食量较大,白天摄食频率高于夜间,摄入的轮虫可布满肠道(图4),镜检发现轮虫壳及其休眠卵无法被鱼苗消化吸收,最终从肛门排出(图5、图6)。
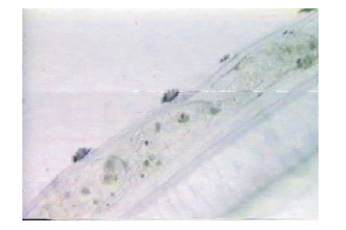
图4 肠道内轮虫
Fig.4 Rotifer in the intestine of the larvae
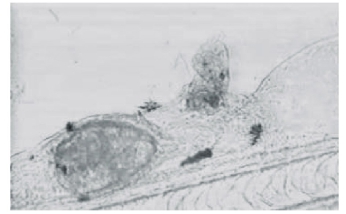
图5 无法消化的轮虫壳从肛门处排出
Fig.5 Shell of rotifers discharging from anus of the larvae
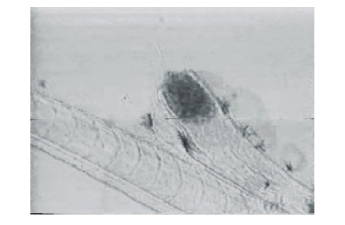
图6 无法消化的休眠卵从肛门处排出
Fig.6 The resting eggs of rotifers discharging from anus of the larvae
本试验中,受精卵从分裂到出膜历时35 d,实际上发育进程主要取决于孵化期总积温,同时还受光照、换水频率的影响[3]。
大银鱼生产性养殖长期以来停滞不前,关键是大银鱼苗种供应无保障。早期仔鱼不肯开口,无法顺利转入主动摄食是主要技术障碍,随着仔鱼卵黄的消耗殆尽即大量死亡。本试验中观察发现,仔鱼出膜后水体中大量卵膜碎屑会在2~3 d内全部自行溶解,试验水体中未进行过人工投饵,但却出现了浮游植物和轮虫,这与养殖水中含有一部分太湖湖水及卵膜碎屑转化为营养物质有一定关系。关于大银鱼仔鱼早期是否摄取浮游植物一直有争议。易
建国等[5]认为,大银鱼仔鱼不能利用藻类。本研究中观察的结果与朱成德[8]报道的大银鱼仔鱼肠道出现少量浮游植物相吻合。荣长宽等[9]亦报道野外水体大银鱼仔鱼对小球藻、衣藻有一定量的摄入。本研究中观察发现,大银鱼仔鱼早期是以浮游植物作为开口饵料(图3),逐步过渡到捕食轮虫(图4)。这也印证了仔鱼在口器的不同发育阶段摄食不同的适口饵料[10]。
关于大银鱼卵膜丝的发生、游离与着生至今尚有争议[11-12],大致可分为:1)卵膜丝从受精孔一极生长出来或粘连于受精孔区,并朝相对一极游离,即“单极正向观点”;2)卵膜丝从受精孔相对的一端发生,向受精孔方向游离,即“单极反向观点”;3)卵膜丝从两极游离出来,形成一环二端呈游离状的细丝,丝中端黏于赤道偏动物极一侧呈圆环状排列,即“两极观点”。本试验结果表明,卵膜丝着生在略偏卵膜孔一侧的赤道环线上,构成环状卵膜丝,这与弋志强等[12]的观察描述相符。但卵膜丝的发源处、游离方向、游离时机以及是单极游离还是两极游离,本试验中尚无充分证据,有待进一步深入研究。
参考文献:
[1] 张开翔.大银鱼胚胎发育的观察[J].湖泊科学,1992,4(2):25 -37.
[2] 龚世园,杨学芬,杨瑞,等.武汉地区大银鱼胚胎发育的观察及其移植[J].华中农业大学学报,1997,16(5):394-398.
[3] 施炜纲,周昕,陈家涤,等.生态因子对大银鱼受精卵孵化和仔鱼存活率的影响[J].水产学报,1998,22(4):375-377.
[4] 郑宝太,李俊杰,姜景田,等.大银鱼池塘养殖技术[J].水产科技情报,2004,31(5):220-221.
[5] 易建国,林长虹.池养条件下大银鱼食性的初步研究[J].水产科技情报,1992,19(5):146-149.
[6] 施炜纲,周昕,孙晓文,等.银鱼移植、增殖技术规范——大银鱼移植、增殖技术[S].北京:中国标准出版社,2002:1-3.
[7] 宋慧春,吴坤明,沈其璋.大银鱼卵膜孔结构的电镜观察[J].动物学报,1999,45(1):8-14.
[8] 朱成德.太湖大银鱼生长与食性的初步研究[J].水产学报, 1985,9(3):275-287.
[9] 荣长宽,岳炳宜.天津大银鱼仔、幼鱼的食性[J].水产科技情报,1998,25(5):204-206.
[10] 尤洋,胡庚东,王博.大银鱼和太湖新银鱼口器结构研究[J].湛江海洋大学学报,1999,19(2):30-33.
[11] 龚世园,张训蒲,杨学芬.大银鱼卵细胞粘丝扫描电镜观察[J].华中农业大学学报,1996,15(6):580-583.
[12] 弋志强,王永玲.大银鱼卵膜表面的扫描电镜观察[J].上海水产大学学报,1998,7(4):348-350.
The embryonic development and habit of larvae in large icefish Protosalartx hyalocranius
SHI Wei-gang,XU Dong-po,LIU Kai,DUAN Jin-rong,ZHANG Min-ying
(Freshwater Fisheries Research Center,Chinese Academy of fishery Sciences,Wuxi 214081,China)
Abstract:The embryonic development and initial feeding of larvae were observed in large icefish Protosalartx hyalocranius under a Pixera Penguin 600CL digital camera system.The fertilized eggs were hatched in 852 hours at water temperature of 4-9℃and the embryonic development was divided into 9 physiological stages,namely 30 developmental substages.The newly hatched larvae had average body length of 4.76 mm and total body length of 4.86 mm.The yolk sac was exhausted about 20 days after hatching,and the fry with body length of 5.85 mm and total body length of 6.09 mm was found to swim normally.The larvae fed on phytoplankton as weaning food in the 10-20 initial feeding days under unfed condition,and began to feed rotifers actively 20-40 days after initial feeding. Microscopic observation revealed that the shells and resting eggs of the rotifers were not digested by the larvae.
Key words:Protosalartx hyalocranius;embryonic development;larva;weaning food
中图分类号:Q954.42
文献标志码:A
文章编号:2095-1388(2011)05-0391-06
收稿日期:2011-01-21
基金项目:公益性行业(农业)科研专项(200903048-03);农业部财政专项(6-125005);中央级公益性科研院所基本科研业务费专项资金(2011JBFC04)
作者简介:施炜纲(1953-),男,研究员。E-mail:shiwg@ffrc.cn