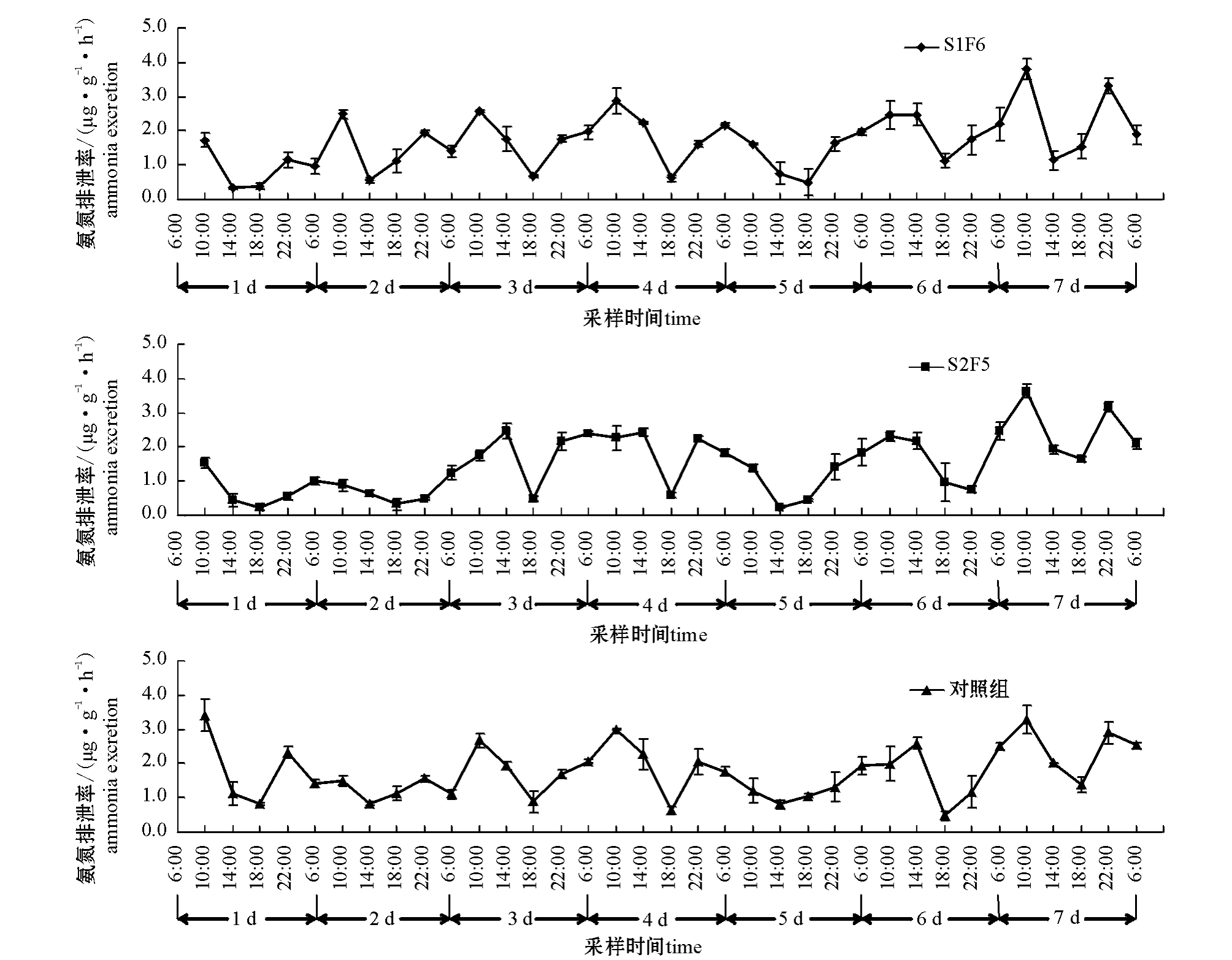
图1 各组牙鲆饥饿及再投喂期间氨氮排泄率的变化(平均值±标准误)
Fig.1 Changes in ammonia excretion rate in Japanese flounder Paralichthys olivaceus during the experiment(Mean±S.E.)
王荦,吴立新,李家庆,吴玉波,陈晶
(大连海洋大学辽宁省水生生物学重点实验室,辽宁大连116023)
摘要:采用静水试验法,在水温为(15.0±1.0)℃下,研究了饥饿及再投喂对牙鲆Paralichthys olivaceus幼鱼(体质量为69.56 g±2.35 g)氨氮、活性磷排泄率的影响。结果表明:饥饿初期牙鲆氨氮、活性磷排泄率呈直线下降,随着饥饿时间的延长排泄率处于相对稳定状态;再投喂后,各饥饿处理组牙鲆氨氮、活性磷排泄率快速恢复到对照组水平,摄食对牙鲆氨氮、活性磷排泄率的影响显著(P<0.05)。在饥饿和饱食状态下,牙鲆氨氮、活性磷排泄率的峰值均出现在10:00,为摄食后4 h,以后逐渐下降;低谷均出现在18:00;再投喂期间平均氨氮排泄率昼夜差异不显著(P>0.05),活性磷排泄率白天略高于夜间(P<0.05)。
关键词:牙鲆;饥饿;再投喂;氨氮;活性磷;排泄率
牙鲆Paralichthys olivaceus是中国重要的海水增养殖鱼类。随着集约化水产养殖业的迅速发展,投饵过剩产生的残饵以及鱼类自身排泄的氮、磷都构成了污染源,这些营养物质随养殖废水的排放进入海区,是导致水体富营养化甚至赤潮的原因之一[1]。排泄是鱼类代谢活动的重要表征,反映了鱼类在外界环境中的生活状况。对鱼类排泄的研究是了解其代谢规律、营养需求和养殖特性等的重要途径[2]。目前,有关鱼类代谢的研究较多[3-8],但尚未见关于饥饿及再投喂对鱼类氮、磷排泄率影响的报道。为此,作者研究了饥饿及再投喂期间牙鲆幼鱼氨氮、活性磷排泄率的变化规律,旨在为优化鱼类投饲制度及合理调控养殖水质提供一些参考。
1.1 材料
试验用牙鲆幼鱼取自大连万洋渔业养殖有限公司,鱼体健壮,无伤病。
1.2 方法
1.2.1 试验鱼的驯养 试验前选取体质量相近的个体放于水族箱中暂养7 d。暂养期间,每天过量投喂牙鲆专用配合饲料(山东升索渔用饲料研究中心生产)2次(6:00和18:00),水温控制在(15.0±1.0)℃,残饵和粪便及时用虹吸管吸出。
1.2.2 试验设计 选取体质量为69.56 g±2.35 g (湿重)的牙鲆幼鱼,采用完全随机分组,设处理组S1F6(饥饿1 d后恢复投喂6 d)、S2F5(饥饿2 d后恢复投喂5 d)、对照组C(试验期间一直饱食投喂),另设水样空白组。每箱放养牙鲆8尾,每组设3个重复,试验共进行7 d。试验期间,每天测定各组水样的氨氮、活性磷5次(测定时间分别为6:00、10:00、14:00、18:00和22:00)。
1.2.3 试验条件及日常管理 试验在实际体积为60 L(60 cm×45 cm×40 cm)的水族箱中进行。试验期间每日投饵两次(6:00和18:00),每次投饵前取样,投喂后及时吸出残饵和粪便。试验用海水经蓄水池沉淀和砂滤处理,投饵后全量换水一次;连续充气,溶氧保持在6 mg/L以上,温度控制在(15.0±1.0)℃,光周期为14L∶10D,盐度为31~33,pH为8.0~8.1。试验采用静水养殖方式。
1.2.4 项目测定及计算 根据《海洋调查规范海水化学要素观测》(GB12763.4—1991)推荐使用的方法,采用次溴酸钠氧化法测定水样中的氨氮,采用抗坏血酸还原的磷钼蓝法测定活性磷酸盐。
鱼类的排泄率E(μg/(g·h))按下式计算:
E=(Ct-C0-△C)V/(t W),
式中:Ct和C0分别为换水前及上一次换水后养殖水体中的氨氮或活性磷酸盐的质量浓度(μg/L);△C为空白组水中氨氮或活性磷酸盐的质量浓度(μg/L);V为试验用水的体积(L);t为排泄试验持续时间(h);W为试验鱼体湿重(g)。
1.3 数据处理
采用SPSS 13.0统计软件进行数据分析,采用单因素方差分析(One-way ANOVA)法检验差异性(P<0.05),用Duncan's法进行多重比较。
2.1 饥饿及再投喂期间牙鲆氨氮和活性磷的排泄率
从图1、图2可见:饥饿期间,牙鲆的氨氮、活性磷排泄率随时间的变化趋势相似。饥饿初期,氨氮、活性磷排泄率呈直线下降;随着饥饿时间的延长,排泄率处于相对较低的稳定状态。再投喂期间,牙鲆的氨氮、活性磷排泄率随时间的变化趋势也相似,呈现出3个阶段:第一阶段,摄食后0~4 h,氨氮、活性磷排泄率迅速上升,达排泄峰值;第二阶段,摄食后4~8 h,排泄率维持在较高水平;第三阶段,摄食后8~12 h,排泄率降到最低。摄食引起氨氮、活性磷排泄率升高的持续时间为4 h,以后排泄率逐渐下降,12 h后达到最低值。
图1 各组牙鲆饥饿及再投喂期间氨氮排泄率的变化(平均值±标准误)
Fig.1 Changes in ammonia excretion rate in Japanese flounder Paralichthys olivaceus during the experiment(Mean±S.E.)
2.2 饥饿及再投喂期间牙鲆的日均氨氮、活性磷的排泄率
从图3、图4可见:饥饿及再投喂期间,各组牙鲆的氨氮、活性磷排泄率峰值均出现在10:00,低谷出现在18:00;再投喂期间平均氨氮排泄率昼夜差异不显著(P>0.05),活性磷排泄率白天略高于夜间(P<0.05)。
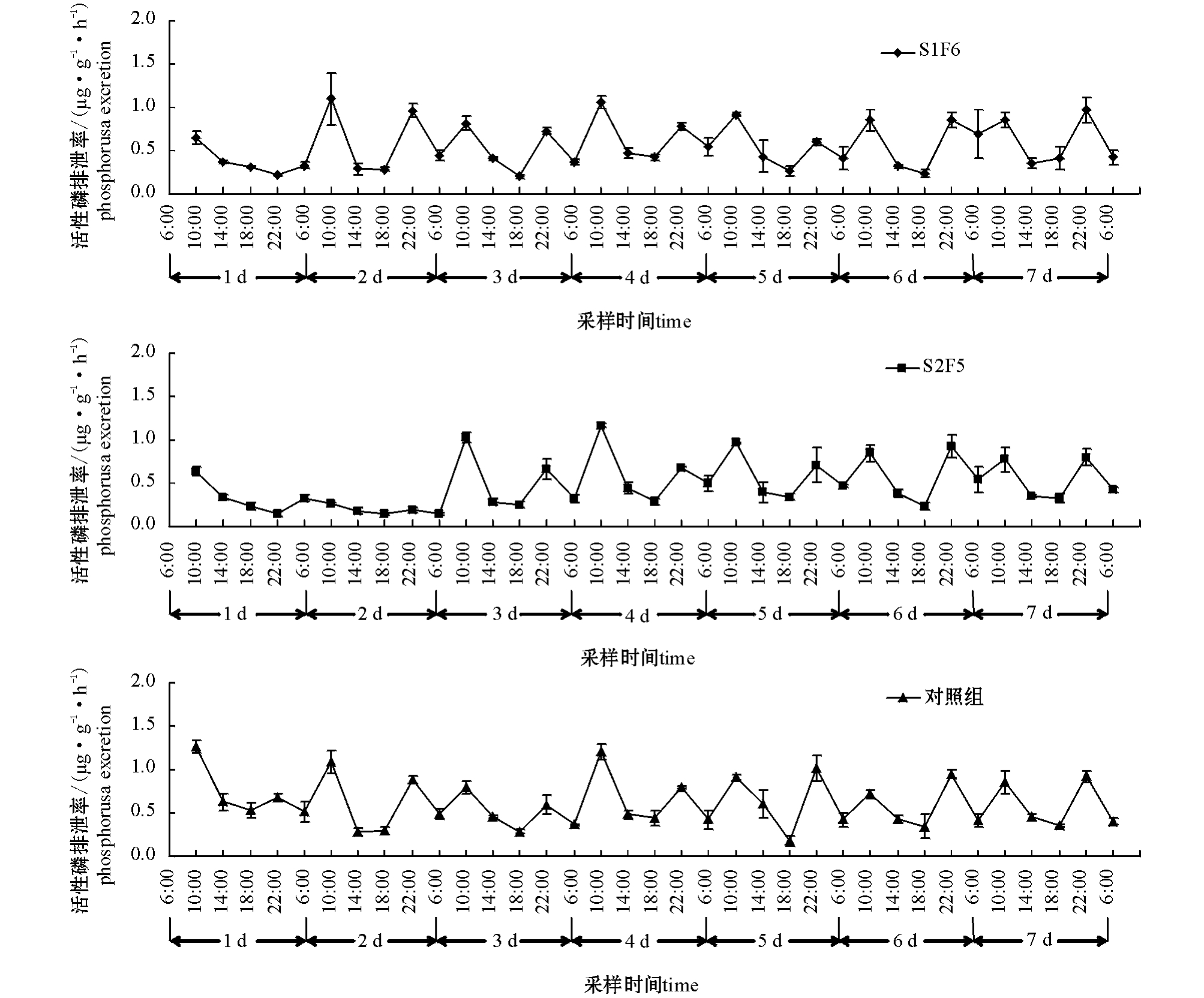
图2 各组牙鲆饥饿及再投喂期间活性磷排泄率的变化(平均值±标准误)
Fig.2 Changes in phosphorus excretion rate in Paralichthys olivaceus during the experiment(Mean±S.E.)
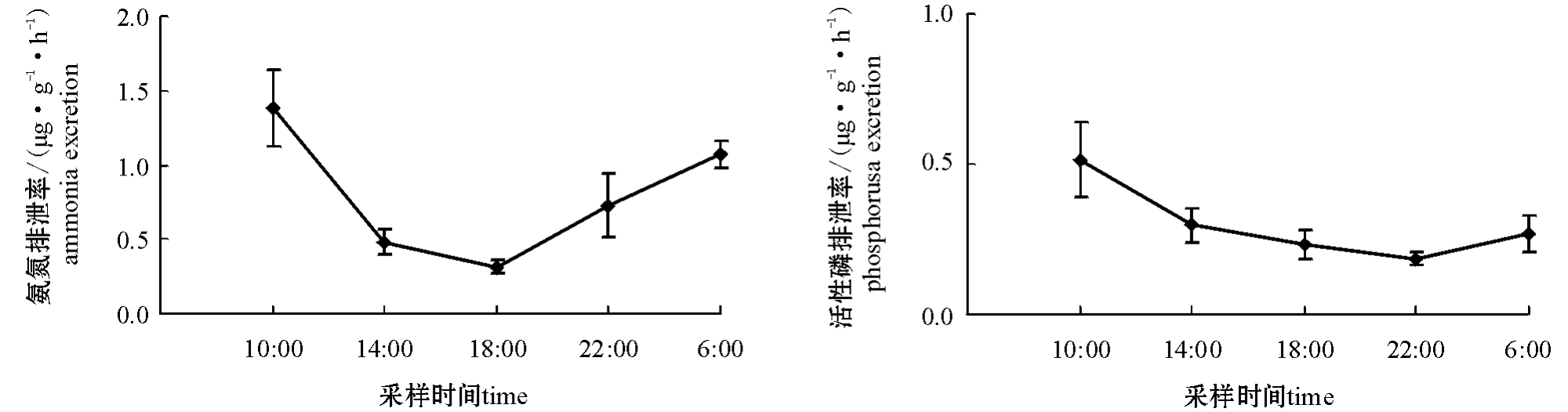
图3 饥饿期间牙鲆日均氨氮、活性磷排泄率的变化(平均值±标准误)
Fig.3 Changes in average daily ammonia and phosphorus excretion rate in Japanese flounder Paralichthys olivaceus during the period of starvation(Mean±S.E.)
3.1 饥饿对牙鲆氨氮、活性磷排泄率的影响
氨和尿素是硬骨鱼类最主要的排泄产物,同时还有极少量的肌酸、尿酸、嘌呤、氨基酸等物质。一般认为,海水硬骨鱼类氨氮占排泄物总量的80%~90%[9],因此,本试验中以测定水样中氨氮的含量作为牙鲆氮的总排泄量。鱼类磷的排泄主要源于动物体蛋白质和脂类的代谢,目前已报道鱼类
所排泄的磷绝大部分都呈磷酸盐形式,某些鱼类的排泄物中磷酸盐占总磷排泄量的85%~95%[10],有的甚至高达100%[11]。因此,本试验中以测定水样中活性磷酸盐的含量作为牙鲆磷的总排泄量。
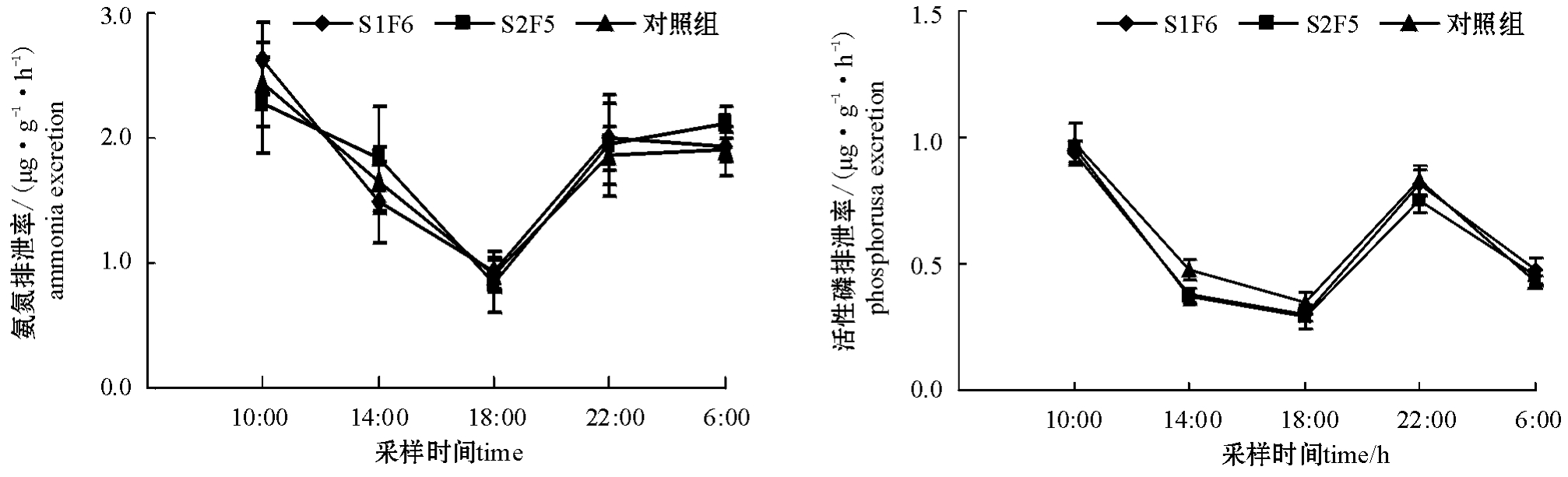
图4 再投喂期间牙鲆日均氨氮、活性磷排泄率的变化(平均值±标准误)
Fig.4 Changes in average daily ammonia and phosphorus excretion rate in Japanese flounder Paralichthys olivaceus during the period of refeeding(Mean±S.E.)
本试验条件下,饥饿期各组牙鲆的氨氮、活性磷排泄率低谷均出现在18:00,峰值均出现在10:00,表明处于饥饿状态下的牙鲆氨氮、活性磷的排泄水平上午可能相对活跃。由此可见,其排泄率的变化呈现一定的阶段性。作者认为,这一规律是牙鲆在饥饿过程中对能量利用适用性要求的综合反映。随着饥饿时间的延长,牙鲆可动用的身体能量不足以维持正常应激状态的能量需求,为了适应环境、延长寿命,牙鲆排泄率很快下降到一个更低的水平,仅保证生存所必须的生理功能所需的能量消耗,并通过调节身体贮存能量的消耗速率使排泄率尽可能保持在较低水平,以便当有食物利用时,可以很快恢复到正常摄食、吸收功能。因为本试验中白天至午夜测定氨氮、活性磷排泄率的时间间隔为4 h,午夜到次日的测定时间间隔为8 h,所以更准确的排泄率峰值和最低值有待于进一步研究。
3.2 再投喂对牙鲆氨氮、活性磷排泄率的影响
再投喂后,各饥饿处理组牙鲆氨氮、活性磷排泄率快速恢复到对照组水平。这可能是由于摄食后鱼体代谢强度增加所致。摄食后的牙鲆主要以蛋白质为代谢物来提供能量,鱼体吸收食物中的氨基酸满足身体需要后多余的部分发生降解,其碳链用以合成储能物质,氨基就被排出而成为氮排泄物。水生动物氨氮排泄量增加的原因比较复杂,一方面可能由于组织中蛋白质的分解代谢所致;另一方面,可能与体内对食物蛋白质进行水解及蛋白质合成有关[12]。通过试验证明,在磷的需求被满足之前,鱼体吸收的所有磷都会在体内累积,不会影响可溶性磷的排泄,然而当鱼体吸收的磷达到其最适需求以后,随着外源磷的摄入,可溶性磷的排泄率将会增加[13]。
已有研究表明,水生动物对食物的消化吸收导致其氨氮排泄量上升,但达到排泄峰值所需要的时间因鱼的种类而异。如红大麻哈鱼Oncorhynchus nerka[14]的氨氮排泄率在摄食后4~4.5 h达到峰值,草鱼Ctenophargnyodon idellus[15]则为5~8 h。排泄率达峰值后逐渐回落至内源性水平,其回落时间受饵料量、食物成分和水温的影响。本试验中饥饿和饱食状态下牙鲆氨氮、活性磷排泄率的峰值均出现在10:00,为摄食后的4 h,以后逐渐下降;低谷均出现在18:00。
研究证实,鱼类代谢水平的昼夜变化规律有3种类型:1)白天代谢水平高于夜间。这些鱼类白天比夜晚进食多、活动较频繁,生命活动及消化吸收需较多的溶氧量,如石鲷Oplegnathus sp.、真鲨Carcharhinus sp.、草鳚Tripterygion minutus和油鲱Brevooria tyrannus、平鲷Rhabdosargus sarba[16]。2)夜间活动较频繁,白天则相对较静止。这些鱼类属于昼伏夜出型,其日均代谢水平低于夜均代谢水平,如六线鱼Hexagrammos sp.和青石斑鱼Epinephalus awoora[17]。3)日均代谢水平与夜均代谢水平大体相似。这些鱼类白天和夜晚都有进食、活动高峰期和低峰期,如大弹涂鱼Boleophthalmus pectinirostris和大西洋牙鲆Paralichthys dentatus[18]。Natarajan[19]、Rosas等[20]认为,昼夜变化是由盐度、温度、摄食和光周期等的变化所控制。而Childress[21]则认为,动物代谢的昼夜变化主要与水体中溶解氧浓度的日变化以及动物为消除溶解氧竞争而采取的生存策略有关。本试验中,氨氮排泄率
昼夜差异不显著(P>0.05),活性磷排泄率白天略高于夜间(P<0.05),这是否与其代谢水平的昼夜变化规律相似,有待于进一步研究。
参考文献:
[1] 计新丽,林小涛,许忠能,等.海水养殖自身污染机制及其对环境的影响[J].海洋环境科学,2000,19(4):66-71.
[2] 童裳亮.鱼类生理学[M].北京:科学出版社,1988:242-243.
[3] 张兆琪,张美昭.牙鲆鱼耗氧率、氮排泄率与体重及温度的关系[J].青岛海洋大学学报,1997,27(4):483-489.
[4] 陈锦云,曹振东,谢小军.温度对瓦氏黄颡鱼幼鱼静止代谢的影响[J].西南师范大学学报:自然科学版,2003,28(4):618-621.
[5] 杨培民,刘刚,张涛,等.鲇幼鱼耗氧率与氨氮排泄率的初步研究[J].大连水产学院学报,2009,24(5):470-474.
[6] 叶安发,周一兵,代智能,等.温度和体重对脉红螺呼吸和排泄的影响[J].大连水产学院学报,2008,23(5):364-369.
[7] 廖志洪,林小涛,王春,等.黄颡鱼仔、稚、幼鱼耗氧率及氨氮排泄率的初步研究[J].生态科学,2004,23(3):223-226.
[8] Lyytikaeinen T,Jobling M.The effect of temperature fluctuations on oxygen consumption and ammonia excretion of underyearling Lake Inari Arctic charr[J].J Fish Biol,1998,52(6):1186-1198.
[9] Dosdat A,Servais F,Metailler R,et al.Comparison of nitrogenous losses in five teleost fish species[J].Aquaculture,1996,141(1-2):107-127.
[10] 阮景荣.三种鱼的磷排泄及其在微型生态系统磷再循环中的作用[J].水生生物学报,2005,29(1):55-60.
[11] Brabrand A,Faafeng B A,Nilssen J P M.Relative importance of phosphorus supply to phytoplankton production:fish excretion versus external loading[J].Can J Fish Aquat Sci,1990,47(2):364-372.
[12] Jobling M.The fish bioenergetics[M].Great Britain:St.Edmundsbury Press,1994:57-64.
[13] Rodehutscord M,Gregus Z,Pfeffer E.Effect of phosphorus intake on faecal and non-faecal phosphorus excretion in rainbow trout Oncorhynchus mykiss and the consequences for comparative phosphorus availability studies[J].Aquaculture,2000,188(3-4):383 -398.
[14] Brett J R,Groves T D D.Physiological Energetics[M].New York:Academic Press,1979:279-352.
[15] 周洪琪,潘兆龙,李世钦,等.摄食和温度对草鱼氮排泄影响的初步研究[J].上海水产大学学报,1999,8(4):293-297.
[16] 丁彦文,李加儿.平鲷Rhabdosargus sarba(Forskal)幼鱼耗氧率的初步研究[J].湛江海洋大学学报,2000,20(3):8-12.
[17] 戴庆年,赵莉莉.青石斑鱼耗氧率的研究[J].水产科学, 1994,13(3):6-9.
[18] 王波,李继强,曹志海.大西洋牙鲆幼鱼标准代谢的初步研究[J].海洋科学进展,2004,22(1):62-68.
[19] Natarajan P.Persistent locomotor rhythmicity in the prawns Penaeus indicus and P.monodon[J].Mar Biol,1989,101(3):339 -346.
[20] Rosas C,Vanegas C,Alcaraz G,et al.Effect of eyestalk ablation on oxygen consumption of Callinectes similis exposed to salinity changes[J].Comp Biochem Physiol,1991,100:75-80.
[21] Childress J J.Effect of pressure,temperature and oxygen on the oxygen-consumption rate of the midwater copepod Gausia princeps [J].Mar Biol,1977,39:19-24.
Effect of starvation and refeeding on excretion of nitrogen and phosphorus in Japanese flounder Paralichthys olivaceus
WANG Luo,WU Li-xin,LI Jia-qing,WU Yu-bo,CHEN Jing
(Key Laboratory of Hydrobiology in Liaoning Province,Dalian Ocean University,Dalian 116023,China)
Abstract:The effect of starvation and refeeding on excretion of nitrogen and phosphorus in Japanese flounder Paralichthys olivaceus(body weight of 69.56 g±2.35 g)were investigated by still water experiment method in a laboratory at water temperature of(15.0±1.0)℃.The results showed that the excretion rate of N and P was sharply decreased in the initial starvation and became relatively stable with the starvation preceeded.After the starved fish were refed,the excretion rates of N and P in the starved fish in the treatment groups were returned to those in the control group,indicating that there were significant effect of refeeding on the excretion of N and P(P<0.05).The maximal N and P excretion rates were found at 10:00 of 4 h after refeeding in the fed and starved fish,and decreased gradually with the minimum at 18:00.There was no significance in diurnal N excretion during the refeeding period(P>0.05).However,the average P excretion was found slightly higher(P<0.05)in the day time than during the night.
Key words:Paralichthys olivaceus;starvation;refeeding;excretion;nitrogen;phosphorus
文章编号:2095-1388(2011)02-0114-05
中图分类号:S967
文献标志码:A
收稿日期:2010-04-26
基金项目:国家科技部“十一五”支撑计划项目(2006BAD09A01);辽宁省教育厅高等学校重点实验室项目(20060180);大连市优秀青年人才基金资助项目(大科技发[2004]166号);辽宁省高等学校优秀人才支持计划项目(RC-05-10)
作者简介:王荦(1985-),女,硕士研究生。E-mail:wang_luoluo@163.com
通信作者:吴立新(1966-),男,教授,博士。E-mail:wulixin@dlou.edu.cn